






外泌體新發現!可抑制宮頸癌鐵死亡!
摘要
誘導癌細胞鐵死亡已被提議作為幾種癌癥類型的潛在治療方法。腫瘤相關巨噬細胞(TAM)在促進腫瘤惡性進展和治療耐藥性方面起關鍵作用。然而,TAM在調節腫瘤鐵死亡中的作用和機制仍未被探索,仍然是個謎。這項研究表明,鐵死亡誘導劑在體外和體內對宮頸癌顯示出治療效果。已發現TAM可以抑制宮頸癌細胞鐵死亡。從機制上講,包裝成外泌體的巨噬細胞衍生的miRNA-660-5p被轉運到癌細胞中。在癌細胞中,miRNA-660-5p減弱ALOX15表達以抑制鐵死亡。此外,巨噬細胞中miRNA-660-5p的上調取決于自分泌IL4/IL13激活的STAT6途徑。重要的是,在臨床宮頸癌病例中,ALOX15與巨噬細胞浸潤呈負相關,這也增加巨噬細胞降低宮頸癌中ALOX15水平的可能性。此外,單因素和多因素Cox分析均顯示ALOX15表達是獨立的預后因素,與宮頸癌的良好預后呈正相關。總之,這項研究揭示靶向TAM在基于鐵死亡的治療和ALOX15作為宮頸癌預后指標中的潛在效用。本文于2023年6月發表在《Acta Pharmaceutica Sinica B》IF:14.5期刊上。
技術路線

主要實驗結果
1、腫瘤相關巨噬細胞抑制宮頸癌鐵死亡
為測試巨噬細胞在調節宮頸癌細胞鐵死亡方面的功能,作者測試用巨噬細胞CM處理的HeLa細胞對鐵死亡誘導劑erastin的敏感性,該誘導劑抑制胱氨酸/谷氨酸抗轉運系統Xc活性。如圖1A和C,通過使用PI染色的細胞死亡檢測,用來自TAM(TAMs-CM)的CM處理的HeLa和SiHa細胞相對于用單核細胞CM(Mo-CM)和M0巨噬細胞CM(M0-CM)處理的細胞對erastin具有更高的抗性。通過使用CCK-8測定來測量用TAMs-CM和三種鐵死亡誘導劑處理的宮頸癌細胞HeLa和SiHa的細胞活力,發現TAMs-CM顯著抑制厄拉斯汀,RSL3(GPX4抑制劑)和索拉非尼(系統Xc抑制劑)在這兩個細胞中的細胞活力降低(圖1B)。
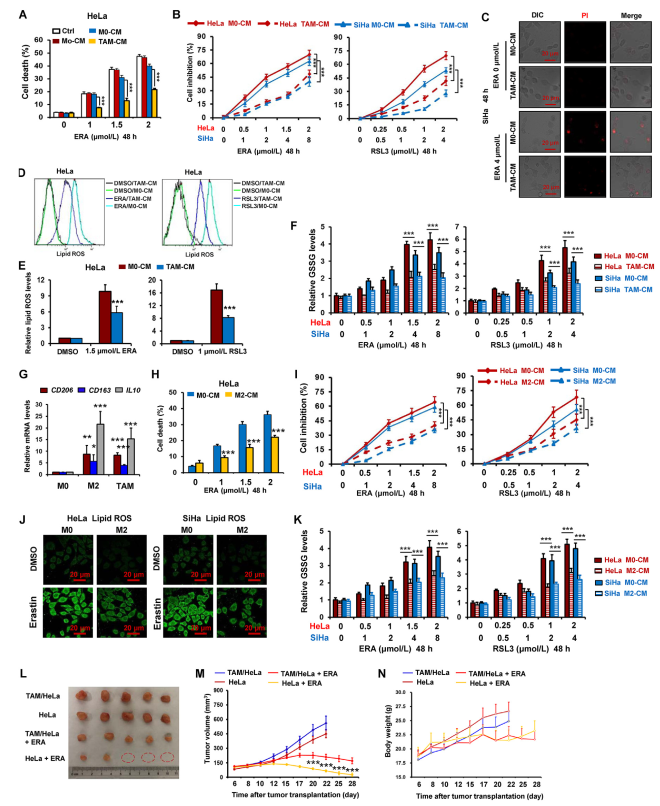
圖1腫瘤相關巨噬細胞抑制宮頸癌的鐵死亡
鑒于脂質過氧化是鐵死亡過程中的重要事件,因此作者研究TAM對erastin和RSL3產生脂質ROS的影響。如圖1D和E所示,TAM顯著抑制HeLa細胞中這兩種藥物引起的脂質ROS的爆發。在索拉非尼處理的HeLa細胞中觀察到相同的結果(圖1)。通過檢測另外兩個鐵死亡標志物MDA和GSSG,作者進一步驗證TAM抑制宮頸癌細胞HeLa和SiHa中的鐵死亡(圖1F)。
先前的研究顯示TAM類似于M2型巨噬細胞。因此,作者探究M2巨噬細胞是否也能減輕宮頸癌細胞的鐵死亡。如圖1G所示,在TAMs和IL4誘導的M2巨噬細胞(M2)中M2巨噬細胞標志物CD206、CD163和IL10的水平明顯升高。圖1H和I表明,由erastin和RSL3誘導的細胞死亡被M2 CM顯著降低。此外,在HeLa和SiHa細胞中,厄拉斯汀和/或RSL3對脂質ROS,GSSG和MDA的誘導顯著減弱(圖1J和K)。
為進一步測試TAMs對體內宮頸癌細胞鐵死亡的影響,作者評估裸鼠HeLa細胞混合TAMs異種移植物中erastin的抗腫瘤活性。如圖1L和M所示,厄拉斯汀顯著抑制腫瘤生長,erastin的抗腫瘤作用被TAMs明顯減弱。此外,在接受erastin治療的小鼠中沒有觀察到明顯的體重損失(圖1N),這表明該藥物在體內的毒副作用很小。
2、TAMs通過外泌體抑制宮頸癌鐵死亡
先前的研究表明,腫瘤微環境中的微囊泡和外泌體促進腫瘤進展和轉移。為探究TAM分泌的外泌體是否增強宮頸癌對鐵死亡的抵抗力,采用超高速離心去除TAMs-CM中的外泌體,然后利用TAMs-CM聯合erastin和RSL3治療HeLa和SiHa細胞。作者推測TAMs衍生的外泌體可以預防鐵死亡。為驗證這一假設,通過離心收集TAMs-CM中的外泌體。外泌體的大小分布和形態分別采用NTA和TEM表征。如圖2A-C所示,外泌體大小異質,TAMs-CM的平均直徑為128±25 nm,M159-CM的平均直徑為21±0 nm。通過蛋白質印跡,作者進一步檢測外泌體的陽性標志物CD63和CD81,以及表征外泌體的陰性標志物Calnexin。圖2D表示CD63和CD81在M0和TAM的外泌體中均高表達,在M0和TAM中表達低。此外,在M0和TAM的外泌體中都沒有Calnexin的表達(圖2D)。
為評估來源于巨噬細胞的外泌體在調節宮頸癌細胞鐵死亡中的功能,分離巨噬細胞外泌體并與erastin處理的HeLa細胞共培養。作者發現,TAMs和M2巨噬細胞來源的外泌體顯著抑制erastin誘導的鐵死亡(圖2E)。此外,通過CCK-8測定,在用TAMs或M2巨噬細胞衍生的外泌體與erastin或RSL3或索拉非尼聯合處理的HeLa和SiHa細胞中測量細胞活力。結果表明,TAM和M2巨噬細胞來源的外泌體都明顯減弱三種鐵死亡誘導劑對細胞活力的降低(圖2F,G)。通過檢測脂質ROS、MDA和GSSG,進一步驗證TAMs來源的外泌體抑制宮頸癌癥細胞HeLa和SiHa中的鐵死亡(圖2H,I)。
為可視化宮頸癌細胞中的外泌體攝取,從TAM中分離外泌體并用CM-DiI標記,然后將HeLa和SiHa細胞與CM-DiI標記的外泌體孵育不同時間。然后,通過流式細胞術檢測外泌體的內化。如圖2J和K所示,具有CM-DiI標記的外泌體的細胞數量以時間依賴性方式增加。通過共聚焦顯微鏡,在0和24小時觀察到內化熒光標記的外泌體。如圖2L和M所示,作者發現HeLa和SiHa細胞都可以有效地吸收外泌體。
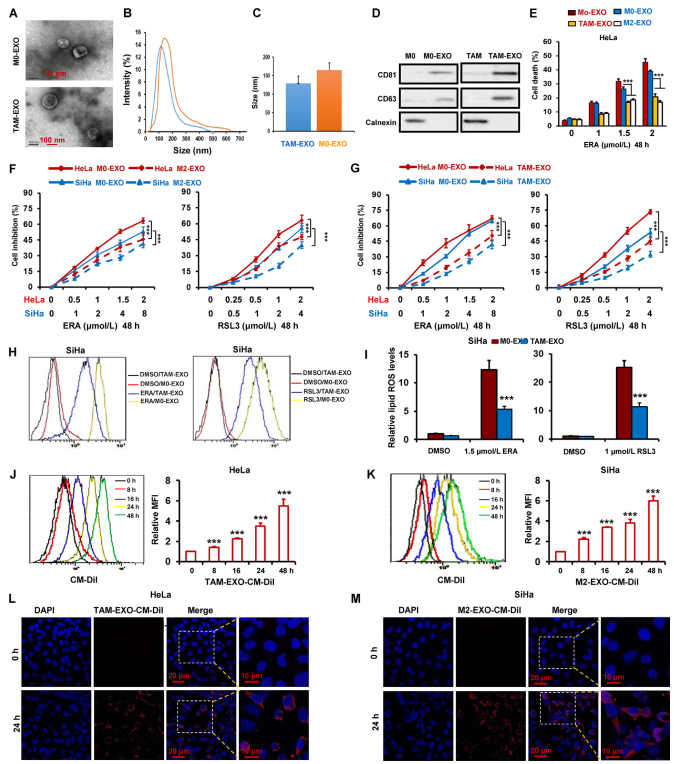
圖2 TAM通過外泌體抑制宮頸癌鐵死亡
3、TAMs對宮頸癌鐵死亡的抑制與ALOX15的下調有關
為探究TAMs抑制鐵死亡的分子機制,作者通過RT-PCR檢測鐵死亡相關基因的mRNA水平,包括NCOA4,BECN1,FTH1,ALOX12,ALOX15,NRF2,FSP1,SCD1,TFRC,LPCAT3,HMGCR,CBS,ATP5G3,CS,P53,ACSL4,GPX4,SLC7A11和LSH。如圖3A所示,通過與M0-CM相比,作者發現TAMs-CM顯著降低HeLa和SiHa細胞中的ALOX15 mRNA水平并略微增加GPX4 mRNA水平。為同時將TAMs-CM改變的基因水平上的倍數變化與全局水平的統計學意義相關聯,以log2倍變化為x軸,log10(P值)為y軸。如圖3B和C所示,火山圖顯示TAMs-CM處理組相對于HeLa和SiHa細胞中M0-CM組的基因表達差異。紅點代表選定的ALOX15基因。這些結果表明,ALOX15是兩個宮頸癌細胞中TAMs-CM最顯著的下調基因。為進一步確認TAMs-CM降低的ALOX15的表達,用PBMC和THP-1衍生的TAM或M2巨噬細胞的條件培養基或純化外泌體處理HeLa細胞。然后,分別通過RT-qPCR和蛋白質印跡檢測ALOX15 mRNA和蛋白質水平。如圖3D-F所示,ALOX15的mRNA和蛋白質水平因條件培養基或TAM或M2巨噬細胞來源的外泌體而顯著降低。在用TAM-CM處理的HeLa細胞中檢測到幾種重要的鐵死亡調節因子FSP1,ACSL4,DHODH和GPX4蛋白表達。
此外,作者在異種移植腫瘤組織中檢測到ALOX15的表達。如圖3G所示,ALOX15在混合TAMs異種移植細胞腫瘤組織中的表達比HeLa細胞異種移植物單獨存在時顯著降低。為檢測ALOX15下調與鐵死亡抑制相關,兩個宮頸癌細胞中ALOX15的表達被siRNA敲低,然后用erastin處理細胞。如圖3H和I所示,敲低ALOX15顯著減少erastin誘導的細胞死亡。為進一步證明ALOX15參與TAMs抑制的宮頸癌細胞鐵死亡,ALOX15被TAMs處理的癌細胞中的siRNA耗盡,然后研究erastin誘導的鐵死亡。如圖3J和K所示,當ALOX15敲除時,TAM不能顯著減弱宮頸癌鐵死亡。
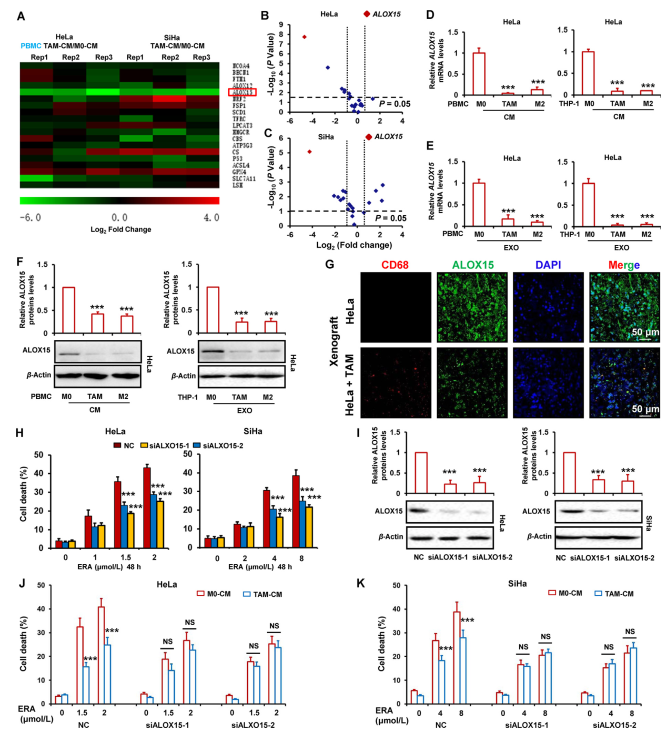
圖3 TAM對宮頸癌癥鐵死亡的抑制與ALOX15的下調有關。
4、TAMs來源的外泌體miR-660-5p減弱ALOX15表達以抑制宮頸癌細胞中的鐵死亡
研究表明巨噬細胞釋放外泌體以將microRNA穿梭到腫瘤微環境中的相鄰癌細胞中。據報道,42個microRNA在M2巨噬細胞來源外泌體中顯著上調,PBMC來源的M2巨噬細胞中有667個microRNA顯著增加。作者從這兩項研究中獲取microRNA的交集,發現10種常見的microRNA(圖4A,B)。接下來,作者將這10個microRNA模擬物轉染到HeLa細胞中并檢測ALOX15表達。如圖4C所示,ALOX15水平被miR-936和miR-660-5p降低。MiR-660-5p對ALOX15表達的抑制作用最顯著。此外,miR-660-5p在TAMs和M2巨噬細胞中的表達明顯高于單核細胞(Mo)和M0巨噬細胞,以及HeLa和SiHa細胞(圖4D)。在TAM、Mo、M0和M2巨噬細胞的外泌體中也檢測到miR-660-5p水平。如圖4D所示,相對于Mo和M0巨噬細胞,TAMs和M2巨噬細胞衍生的外泌體中miR-660-5p水平顯著增加。
為研究miR-660-5p在介導TAMs抑制宮頸癌癥細胞ALOX15表達中的作用,用miR-660-5p抑制劑轉染HeLa細胞,然后用M0巨噬細胞和TAMs的CM和外泌體處理。通過RT-qPCR和蛋白印跡分別檢測ALOX15的mRNA和蛋白水平。圖4E和F表明,相對于對照M0巨噬細胞,ALOX15的表達被TAMs的CM或外泌體顯著抑制,而miR-660-5p抑制劑明顯逆轉CM或TAMs來源外泌體對ALOX15表達的減少。
由于microRNA主要通過與基因的mRNA結合來減弱基因表達,因此作者探究miR-660-5p是否與ALOX15基因的mRNA結合。利用在線miRNA靶生物信息學預測數據庫miRWalk預測ALOX660編碼序列(CDS)中保守的miR-660-5p靶位點(圖4G)。為進一步確認miR-660-5p對ALOX6605的靶向,進行熒光素酶活性測定。將ALOX15的野生型(WT)或突變型(MT)CDS克隆到pGL3熒光素酶報告載體中螢火蟲熒光素酶編碼區的下游。載體與miR-660-5p模擬物或抑制劑共轉染到HeLa細胞中。正如預期的那樣,作者發現miR-660-5p模擬物急劇降低熒光素酶活性,而miR-660-5p抑制劑顯著增加熒光素酶活性(圖4H)。此外,在轉染MT報告載體的HeLa細胞中,miR-660-5p模擬物和抑制劑對熒光素酶活性沒有明顯變化(圖4H)。在HeLa和SiHa細胞中,作者也顯示出模擬物和抑制劑可以明顯降低和誘導ALOX15 mRNA表達(圖4I)。用熒光素酶報告載體轉染的HeLa細胞與ALOX15的WT或MT CDS進一步用TAM-和M2巨噬細胞-CM處理,然后檢測熒光素酶活性。圖4J表明TAMs和M2巨噬細胞-CM 顯著下調WT CDS轉染HeLa細胞報告載體中的熒光素酶活性,但不下調MT CDS。
為研究miR-660-5p在TAMs抑制的鐵死亡中的作用,用miR-660-5p抑制劑轉染HeLa和SiHa細胞,并與TAM的CM和外泌體一起孵育。然后用erastin和RSL3處理這些細胞。通過檢測細胞死亡,作者發現CM和外泌體對細胞死亡的減少被miR-660-5p抑制劑完全阻斷(圖4K和L)。這一結果表明,宮頸癌細胞中內源性miRNA-660-5p的表達太低,無法影響細胞鐵死亡反應。
此外,采用腫瘤異種移植裸鼠模型評估miR-660-5p在介導TAMs誘導的宮頸癌體內鐵死亡抵抗中的作用。如圖4M,在厄拉斯汀治療后,異種移植腫瘤與miR-660-5p抑制劑高表達HeLa細胞和TAMs聯合生長比TAMs和對照HeLa的腫瘤生長更快,更大。然而,在那些單獨使用HeLa的腫瘤中,miR-660-5p抑制劑未能改變腫瘤對erastin的敏感性。
為直接觀察TAMs衍生的miRNA進入腫瘤細胞,將HeLa和SiHa細胞與在宮頸癌細胞中轉染FAM標記的miR-660-5p模擬物的TAM共培養(圖4P),并通過熒光顯微鏡觀察。圖4N顯示HeLa和SiHa細胞中都存在FAM熒光。此外,流式細胞術分析顯示,用FAM標記的microRNA轉染的兩個癌細胞(圖4O).
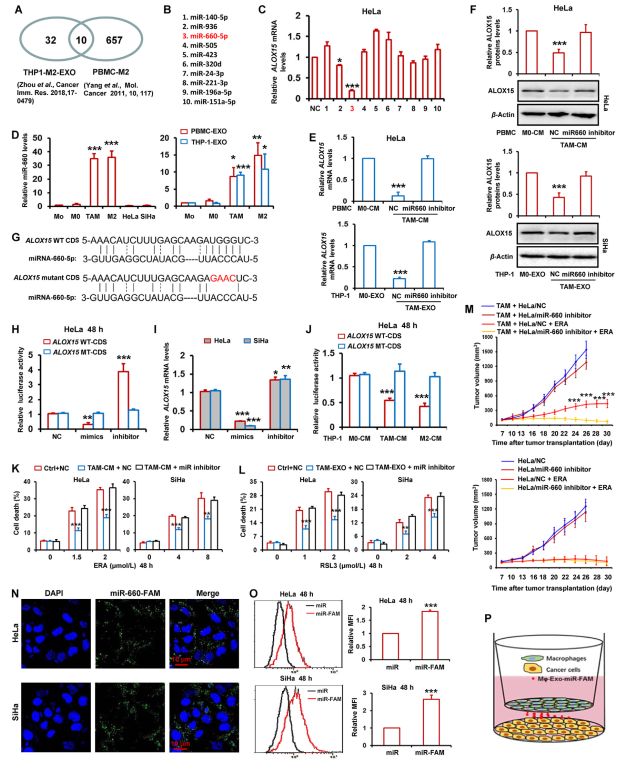
圖4 TAMs來源的外體miR-660-5p減弱ALOX15的表達,以抑制宮頸癌細胞鐵死亡
5、IL4/IL13通過激活STAT660途徑誘導miR-5-6p表達
作者發現miR-660-5p在TAM和M2巨噬細胞中均過表達。據報道,TAM的許多特性也是M2巨噬細胞的特征。幾種細胞因子在兩種細胞中均有表達,如IL6、IL10、IL4、IL13和TGF-β。因此,作者想知道這些因素的表達是否與miR-660-5p有關。通過RT-qPCR測定,作者發現在TAMs中,IL4和IL13以劑量依賴的方式增加miR-660-5p的水平(圖5A),而IL6,IL10和TGF-β不影響miR-660-5p水平(圖5B和C)。先前的研究表明,STAT3和STAT6激活是IL4和IL13的原因。因此,作者假設TAM中IL3和IL6激活的STAT4和STAT13與miR-660-5p表達相關。通過抑制TAM中STAT3和STAT6的活化,作者發現IL660和IL5對miR-4-13p的誘導被STAT6抑制劑而不是STAT3抑制劑顯著減弱(圖5D和F)。蛋白質印跡結果顯示,兩種抑制劑分別顯著降低p-STAT6和p-STAT3(即活化的STAT6和STAT3)(圖5E和G)。圖5H還表明IL4和IL13均顯著增加TAMs外泌體中的miR-660-5p水平,而IL660和IL5誘導的miR-4-13p水平明顯減弱。通過檢測巨噬細胞中的miR-660-5p和p-STAT6表達,作者發現miR-660-5p和p-STAT6在TAMs和M2巨噬細胞中的水平明顯高于M0巨噬細胞(圖5I-L)。為研究STAT660抑制劑對miR-5-6p表達的抑制是否減弱TAM誘導的鐵死亡抵抗力,將帶有Hela混合TAM的異種移植腫瘤與STAT6抑制劑和erastin聯合治療。如圖5M所示,STAT6抑制劑顯著減弱TAM誘導的對erastin的耐藥性。然而,在單獨使用Hela的異種移植腫瘤中,作者發現STAT6抑制劑還增強erastin(圖5N)。在兩種腫瘤模型中,單獨使用STAT6抑制劑發揮一定的抗腫瘤作用(圖5M和N)。這表明STAT6抑制劑增強erastin-抗腫瘤作用可能參與TAM中miR-660-5p表達的抑制和腫瘤細胞中STAT6活性的抑制。此外,注射erastin和STAT6抑制劑的小鼠體重沒有顯著變化(圖5O和P)。
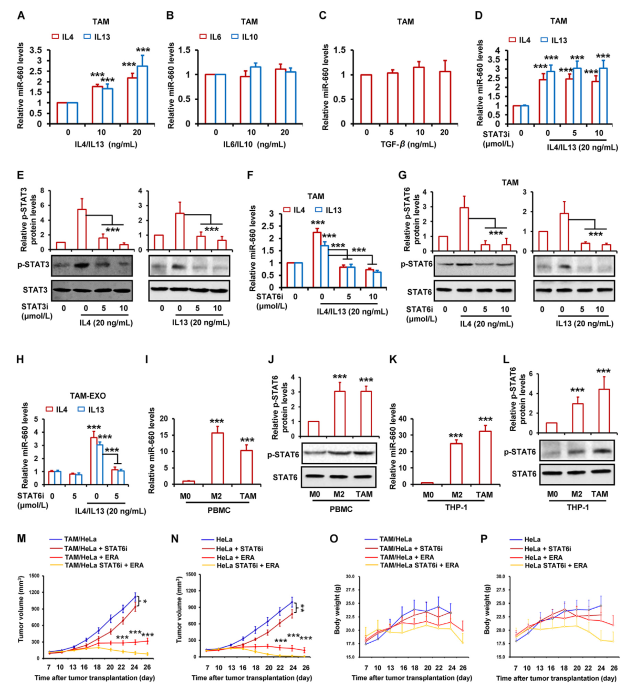
圖5 IL4/IL13通過激活STAT6途徑誘導miR-660-5p表達
6、ALOX15與巨噬細胞浸潤呈負相關,與良好預后呈正相關
為進一步證實TAMs抑制宮頸癌中的ALOX15表達,分析TCGA中高表達與低表達ALOX15宮頸癌組織中免疫細胞群的豐度。圖6A顯示高表達ALOX15宮頸癌組織中巨噬細胞豐度顯著降低。此外,作者通過熱圖來可視化28個浸潤免疫細胞群(圖6B)。結果表明,與高表達的ALOX15組織相比,低表達ALOX15宮頸癌組織中巨噬細胞的標準化ssGSEA評分顯著更高。上述結果提出浸潤TAMs抑制宮頸癌細胞ALOX15表達的可能性。
此外,ALOX15表達與TCGA中宮頸癌癥OS和無病生存率呈正相關(圖6C和D)。此外,鄭州大學進一步分析ALOX15表達與宮頸癌預后的關系。通過使用IHC染色,研究ALOX15在147個石蠟包埋的存檔宮頸癌組織中的表達。組織包括53個Ib1期,28個Ib2期,31個IIa1期和35個IIa2期腫瘤。在147個樣品中,在82個樣品中檢測到ALOX15蛋白的高表達,在其余65個樣品中觀察到ALOX15的低水平(表1和圖6E)。患者生存率分析顯示,宮頸癌癥患者的ALOX15表達與OS之間存在明顯的正相關性(圖6F)。
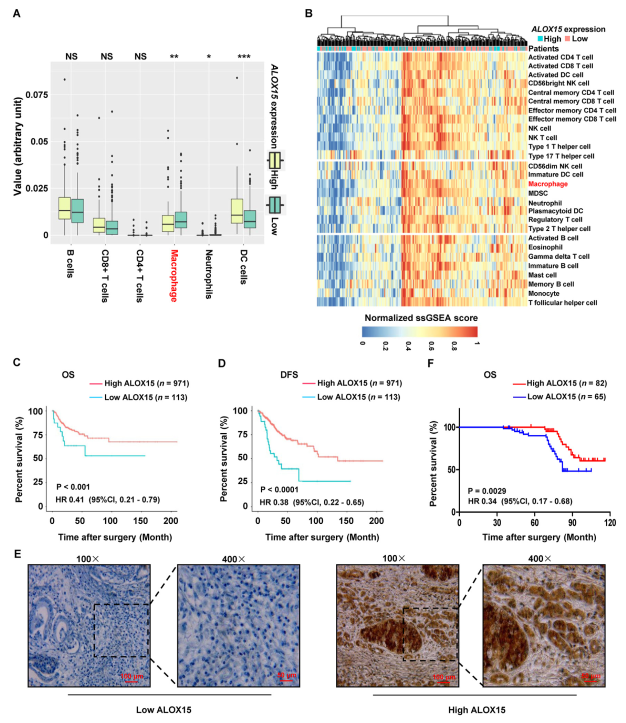
圖6 ALOX15與巨噬細胞浸潤和良好預后有關
根據宮頸癌患者IHC評分,分析ALOX15水平與臨床病理特征的相關性。如表1,高ALOX15表達與腫瘤大小、復發率和5年內生命狀態呈顯著負相關。然而,其他不良的臨床病理特征,如年齡和FIGO分期,與ALOX15表達沒有顯著相關性。為驗證ALOX15表達與宮頸癌預后之間的關系,作者進行單變量和多變量Cox分析。單因素分析顯示,腫瘤大小、淋巴血管間隙受累、T分類、盆腔淋巴結轉移和ALOX15表達是OS的預后因素(表2)。多因素分析顯示,淋巴血管間隙受累、T分類和ALOX15表達是獨立的預后因素(表2)。
最后,該研究提供一個TAM增強宮頸癌對鐵死亡抗性的示意圖(圖7)。
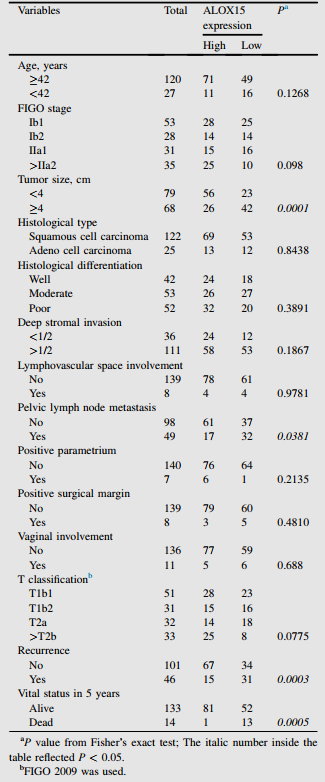
表1 ALOX15表達與宮頸癌臨床病理特征的相關性
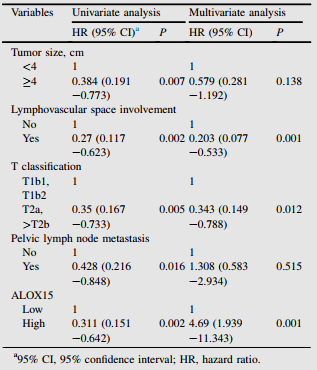
表2 癌癥臨床病理特征與總生存率相關的單變量多變量Cox回歸分析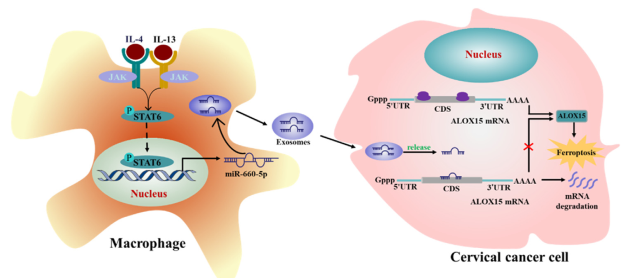
圖7 TAM增強癌癥對鐵死亡抗性的示意圖。一個說明TAM來源的外泌體miR-660-5p在調節ALOX15誘導的宮頸癌癥細胞鐵死亡中作用的假說模型。
總之,該研究表明TAMs通過分泌外泌體傳遞特殊信號分子來抑制宮頸癌細胞鐵死亡的新功能。研究還為TME抑制的癌細胞死亡機制提供新的見解。此外,STAT6抑制劑增強鐵死亡誘導劑的抗腫瘤作用,表明STAT6抑制劑與鐵死亡誘導劑的聯合應用可能是宮頸癌新的有前景的治療方法。同時發現ALOX15表達與患者總生存期和無病生存期呈顯著負相關,表明ALOX15可能是宮頸癌的預后標志物。
實驗方法
細胞活力和細胞死亡測定,脂質ROS分析,丙二醛(MDA)和谷胱甘肽的測定,RNA干擾,雙熒光素酶活性測定,RT-qPCR,WB,免疫熒光,外泌體純化和標記,小鼠異種移植宮頸癌模型,透射電子顯微鏡(TEM)測定,免疫組織化學(IHC)分析
參考文獻
Luo Y, Chen Y, Jin H, et al. The suppression of cervical cancer ferroptosis by macrophages: The attenuation of ALOX15 in cancer cells by macrophages-derived exosomes. Acta Pharm Sin B. 2023; 13(6): 2645-2662. doi:10.1016/j.apsb.2023.03.025