






揭秘肝臟p53依賴的組織特異性抗輻射機制
胸腺和脾臟與肝臟不同,是對輻射敏感的組織,在體內全身輻射后會觸發p53依賴性凋亡。但這種肝臟特異性抗性的分子機制尚不清楚。作者通過對經輻射處理的小鼠器官進行RNA-seq和ChIP-seq聯合分析,發現了共有的和組織特異的p53轉錄反應。胸腺和脾臟共有的p53靶點富含凋亡靶點。肝臟不能上調這些基因并不是由于基因占有率降低。p53的C末端缺失可使輻射誘導的p53凋亡靶點在肝臟表達,同時增加細胞死亡。肝臟特異性p53靶標的轉錄激活還需要C末端的額外作用。據作者推測,抑制凋亡基因的表達與增強肝臟特異性靶點的活化共同賦予了組織特異性抗輻射能力。本文于2023年3月發表在《Cell Reports》IF:8.8期刊。
技術路線

主要實驗結果
1、DNA損傷可誘導小鼠胸腺和脾臟p53依賴性細胞凋亡,但對肝臟無影響
以前的研究發現胸腺和脾臟是體內輻射敏感組織,而肝臟不是。為驗證這些結論,對21天大的野生型C57/BL6小鼠進行2Gy的全身X射線照射。在處理6小時后,胸腺和脾臟出現大量的凋亡,而肝臟則沒有(圖1A)。使用p53基因缺失的小鼠證明胸腺和脾臟的這些效應確實是p53依賴性的(圖1B)。免疫印跡法用于確定三種組織中p53的相對表達水平。在這些研究中,凝膠中的每個泳道代表一只動物。在X射線照射3小時后,p53在三種組織中的表達被誘導,并在6小時后開始減弱(圖1C)。定量測量p53和actin條帶的相對強度表明,盡管肝臟p53的表達水平可能較低,但無論是在治療前(0 h)還是在輻射后(3 h),這三種組織中p53的表達水平沒有統計學意義上的顯著差異(圖1D)。仔細研究p53蛋白水平對輻射的反應表明,胸腺和脾臟中的p53有統計學意義的增加(分別為p = 0.002和p = 0.001),但肝臟中的情況并非如此(p = 0.36)。然而,肝臟顯示出強大的p53靶基因反應(見下文)。
2、X射線處理導致共同的和組織特異性的p53轉錄反應
為確定對全局基因表達的影響,野生型和p53缺失小鼠均未接受或接受2 Gy的X射線照射。從肝臟、胸腺和脾臟中收集RNA,并進行RNA-seq分析。基因表達顯著改變的基因數量因組織而異,脾臟最多(3132個基因),其次是胸腺(481個基因)和肝臟(324個基因)。在這三種組織中,基因表達的上調高于抑制(圖1E)。為確定這些基因表達變化對p53的依賴性,在相應的p53-null組織中測定輻射對野生型組織中顯著改變的基因表達的影響(圖1F)。火山圖顯示,在肝臟中明顯改變的324個基因中,298個(92%)的表達依賴于p53。有趣的是,所有胸腺基因的表達都依賴于p53。但在脾臟中,在3132個已鑒定的基因中,298個基因在p53缺失小鼠中也發生了改變,其中2834個基因(90%)依賴于p53(圖1F)。
對相應組織進行染色質免疫沉淀測序(ChIP-seq)分析,以確定p53的占據位點。基因表達水平脾臟顯示了最多的輻射改變基因(3132個,而肝臟為324個,胸腺為481個)(圖1E),而與此相反,肝臟具有最多的p53基因占據的重要位點(3872個,而胸腺為2405個,脾臟為2604個)(圖1G)。將RNA-seq數據與ChIP-seq數據進行整合,以鑒定對輻射反應顯著表達改變同時也有相關的ChIP-seq峰的基因。這樣就鑒定出在放射后上調(圖1H)或下調(圖1I)的真正的p53靶點。盡管上調基因在不同組織間有相當大的重疊,但下調靶點具有高度的組織特異性。

圖1 DNA損傷誘導胸腺和脾臟p53依賴性細胞凋亡,而不是肝臟,其具有共同的和組織特異性的p53轉錄反應
3、X射線誘導了p53的共享核心直接轉錄靶點
整合RNA-seq和ChIP-seq分析發現,20個基因在肝臟、胸腺和脾臟這三種組織中均顯著上調,且在基因的10 kb范圍內或基因本身至少有一個ChIP峰。這20個基因的上調在所有受檢組織中都依賴于p53(圖2A和2B)。選擇了三個基因進行qRT-PCR驗證(圖2C)。Ccng1、Phlda3和Aen都上調了3-12倍,前兩個基因在每個組織中都明顯依賴于p53(圖2C)。對這三個基因的ChIP-seq圖譜的研究證實,在靠近轉錄起始位點的地方存在穩健的峰,且在Phlda3中,在基因上游存在額外的峰(圖2D)。在這20個基因中,所有基因都至少有一個峰與p53 motif相關(圖S3B),這些motif與之前發表的p53特異性DNA結合共識序列非常吻合(圖2D),對所有20個共有基因的類似分析提供了一個與之前確定的非常相似的motif(圖2E)。有趣的是,這些峰值在基因體內本身也同樣存在,如在基因的上游,只有少數位點位于下游(圖2F)。GSEA顯示,GO_BP與先前與導致細胞凋亡的p53 DNA損傷應答相關的基因有關(圖2G)。綜上所述,20個共享基因的發現(其中許多基因先前已被鑒定為p53靶標)使人們對這些RNA-seq和ChIP-seq數據集及其用于進一步評估充滿信心。
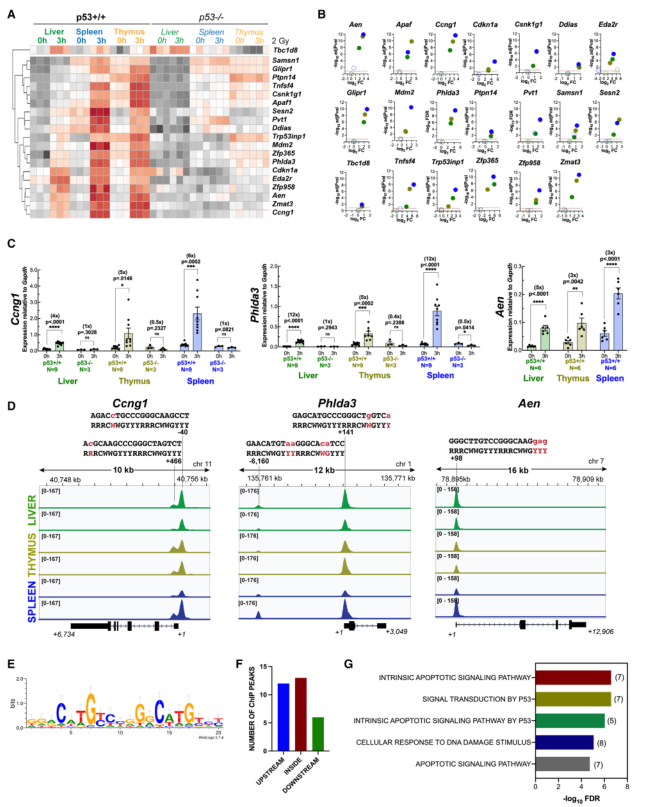
圖2 X射線誘導了p53的共享核心直接轉錄靶點
4、肝臟p53不能上調凋亡基因
為闡明胸腺和脾臟與肝臟對輻射的不同生物反應的基礎,接下來確定了敏感組織中特異性上調的基因。結果發現,80個基因在胸腺和脾臟中的表達在輻射后增加,而在肝臟中沒有變化(圖3A)。所有這80個基因都有與之相關的ChIP峰,并被認為是p53的真正靶標。GSEA顯示與細胞凋亡相關的生物過程是富集率最高的(圖3B)。與凋亡過程基因集重疊的21個基因被證實其基因表達依賴于p53,并且在肝臟中不表達(圖3C)。qRT-PCR被用來證實這些基因的一個子集(Bbc3、Pmaip1和Bax)在胸腺和脾臟中確實是選擇性靶點,但在肝臟中不是,并且它們的表達依賴于p53(圖3D)。一個共同的靶點Cdkn1a在所有三個組織中作為p53依賴性基因表達的陽性對照。在10周齡小鼠中檢測Bbc3 (Puma)時,在蛋白和mRNA水平發現了類似的組織特異性。值得注意的是,與胸腺和脾臟相比,肝臟的DNA損傷反應減弱,這可通過免疫印跡法檢測p53在絲氨酸15處的磷酸化和γH2AX的水平來確定(圖S2)。這就提出了一種可能性,即基因表達的組織特異性可能與對X射線的不同敏感性有關。盡管如此,肝臟確實顯示出其他組織所沒有的基因亞群的占據和表達(圖1H)。肝臟特異性p53靶點的存在降低了上游信號傳導受阻導致觀察到的結果的可能性,但并不排除這種可能性。
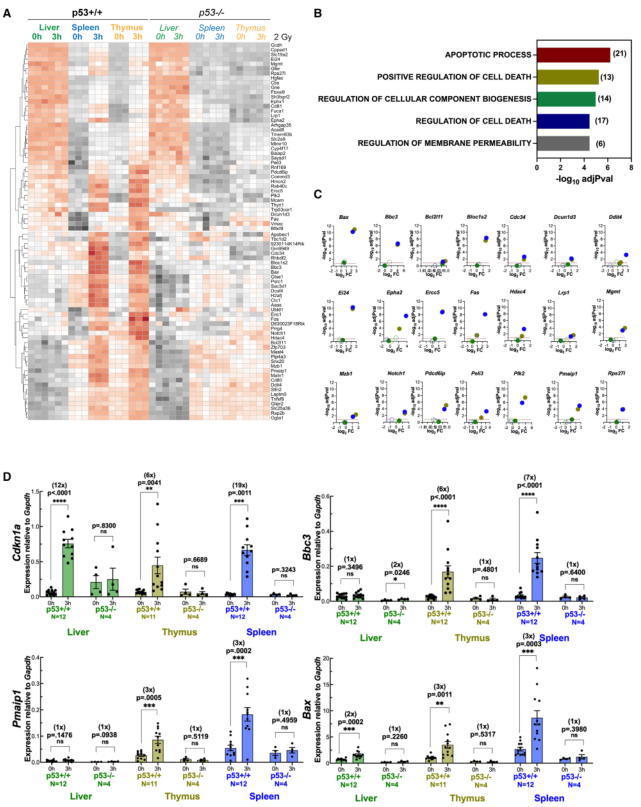
圖3肝臟p53不能上調凋亡基因
5、無法上調肝臟中的p53凋亡靶點不是由于基因占用減少
肝臟p53不能上調凋亡靶點的分子基礎可能是由于基因占據受損或DNA結合后的步驟。為解決這個問題,作者研究了三個關鍵凋亡靶標(Bbc3、Pmaip1和Bax)的ChIP-seq圖譜。在輻照胸腺和脾臟中,三個基因都觀察到強大的p53峰值。令人驚訝的是,在肝臟中,這三個基因的p53占據率也相似(圖4A)。為了驗證這一發現,野生型p53小鼠體內用另一種DNA損傷劑多柔比星處理,并對肝臟和脾臟組織進行分析。進行qRT-PCR檢測兩個共享靶標Cdkn1a和Mdm2的mRNA水平,并與兩個凋亡靶標Bbc3和Pmaip1進行比較。與X-射線照射后的結果一致,多柔比星處理導致肝臟和脾臟中Cdkn1a和Mdm2的增加,而Bbc3和Pmaip1僅在脾臟中被誘導(圖4B)。然后對相應的肝臟和脾臟組織樣本進行染色質免疫沉淀。與X-射線的結果一樣,多柔比星處理后,在肝臟和脾臟的Cdkn1a、Bbc3和Mdm2基因的反應元件上檢測到p53的占據,Alb基因的一個區域被用作陰性對照(圖4C)。
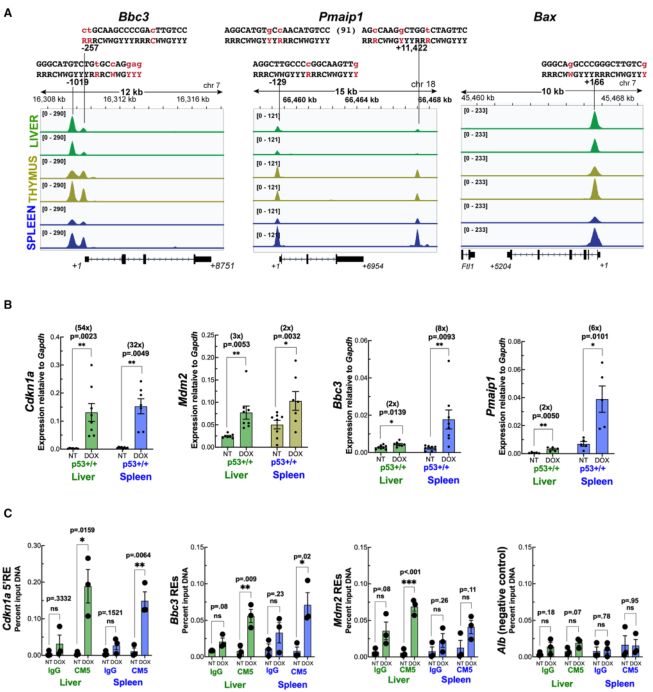
圖4無法上調肝臟中的p53凋亡靶點不是由于基因占用減少
6、肝臟中p53靶基因表達的抑制有兩種機制
迄今為止的研究結果表明,胸腺和脾臟靶標中的三個,即Bbc3、Pmaip1和Bax,被肝臟p53占據,但在該組織中對X射線或多柔比星處理沒有調節(圖3和圖4)。然后研究了80個胸腺和脾臟靶點中有多少具有類似的特性,即它們也被p53占據但在肝臟中不被上調。在胸腺和脾臟中,所有這80個基因都顯示出與相應的p53占位有顯著差異的基因表達(圖5A)。有趣的是,這些基因中的大多數在肝臟中也顯示出顯著的占位,調整后的p < 0.20(圖5A)。值得注意的是,與胸腺和脾臟相比,這些基因在肝臟中的占據率在統計學上更高(圖5B,左)。兩個代表性基因(Notch1, Ei24)的ChIP-seq圖譜顯示,p53在所有三個組織中都有很強的占據率(圖5B,中)。總之,80個基因中有74個被肝臟p53占據,其中所有與GO凋亡過程基因組重疊的基因都在這里被發現(下劃線基因,圖5B,右)。這些峰值主要出現在上游區域(圖5B,左下)。將這些峰中的p53 motif進行比對,發現它們與之前確定的p53共識有合理的匹配(圖5B,右下)。
相比之下,其余6個基因在肝臟中沒有與之相關的穩健的p53 ChIP峰(圖5C,左)。展示了ChIP-seq圖譜的兩個例子(Hgfac和Dcaf4)(圖5C中)。圖5C右展示了6個基因的列表。在胸腺和脾臟中發現的這些峰值,但在肝臟中沒有發現,也大多出現在基因體內(圖5C,左下)。這些峰中的p53 motif也與p53共識相似(圖5C右下)。
值得注意的是,在74個顯示肝臟p53占位的基因中,有7個基因有多個相關的ChIP峰,其中至少有一個峰在肝臟樣本中丟失。圖5B右側紫色顯示了這些基因。在ChIP峰與特定p53靶基因的功能注釋出現之前,很難明確論證這一點的相關性。然而,肝臟p53對這些基因缺乏調控仍有可能是由于這些其他位點缺乏占位。
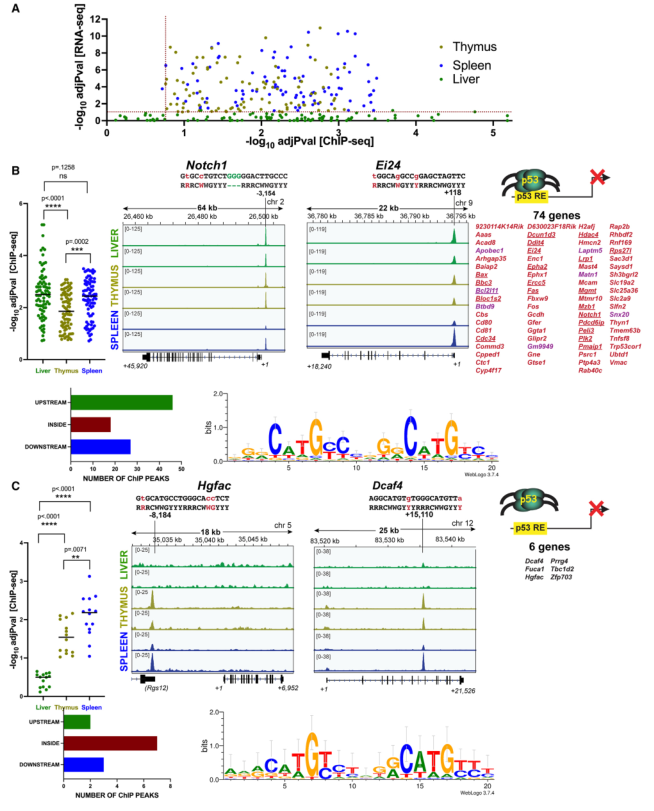
圖5 肝臟p53靶基因表達缺陷有兩種機制
7、p53的C端阻止了肝臟中p53凋亡靶點的上調
肝臟p53不能上調胸腺和脾臟的大部分靶標似乎不是由于基因占據受損。這表明下游轉錄事件受到了影響,這種影響很可能是由于蛋白質與蛋白質之間的相互作用。由于p53的CTD過去已被證實可介導p53與基因表達相關的輔助因子的結合,因此研究了受損肝臟p53反應對CTD的需求。之前文獻已經產生了一個小鼠模型,其中內源性Trp53基因表達了一個缺少最后24個氨基酸的截短蛋白。同卵p53Δ24/Δ24小鼠在出生后兩周內死亡,因此不適合在此進行研究。然而,雜合子p53Δ24/-小鼠壽命正常,無明顯表型。因此,對p53Δ24/-小鼠進行X-射線處理并分析p53靶點的表達。與p53+/-小鼠相比,Δ24/-小鼠肝臟、胸腺和脾臟中某些p53靶點的表達進行了檢測(圖6)。CTD的缺失對胸腺或脾臟中共有靶點Cdkn1a和Mdm2或胸腺/脾臟特異靶點Bbc3、Pmaip1和Bax的基因表達沒有實質性影響(圖6A)。相反,缺失CTD后,肝臟中Cdkn1a、Bbc3、Pmaip1和Mdm2的誘導作用增強,但Bax的誘導作用不增強(圖6A)。值得注意的是,輻射后p53+/-肝臟中mRNA幾乎沒有變化的Bbc3,現在在p53Δ24/-肝臟中被輻射誘導。進行免疫印跡以確定CTD的缺失是否改變了p53蛋白的表達(圖6B)。多個印跡的定量證實不同組織的p53水平相當(圖6C)。p53蛋白水平的誘導在不同組織和基因型中同樣相似。與p53Δ24/-肝臟中Bbc3 mRNA的誘導一致,Puma蛋白也被強有力地誘導,并具有統計學意義(圖6B和6C)。因此,CTD的缺失允許肝臟特異性上調凋亡靶標,特別是在輻射處理后的Bbc3。

圖6 p53的C端阻止了肝臟中p53凋亡靶點的上調
8、p53的C端阻止X射線誘導的肝臟凋亡
為觀察CTD缺失后觀察到的基因表達變化是否會影響X射線治療后的生物學結果,對p53+/-和p53Δ24/-小鼠的肝組織進行了TUNEL檢測(圖7A)。顯微照片證實p53+/-肝臟在輻射前后幾乎沒有TUNEL陽性(圖7A,左)。這與同源野生型p53小鼠的情況相似(圖1)。相反,p5324/-肝臟顯示TUNEL陽性細胞增加。對多個實驗中的此類顯微照片進行定量顯示增加了7倍(圖7A,右)。這與CTD缺失增強肝臟輻射敏感性的觀點一致,同時伴隨著關鍵凋亡靶標Bbc3及其相應蛋白Puma表達的增加。鑒于全長p53在這些組織中的基礎凋亡率很高,因此與肝臟相比,去除CTD的影響輕微也就不足為奇了(圖7A)。
9、p53的C端是肝臟特異性基因表達的關鍵決定因素
CTD缺失對輻射誘導的在肝臟中的一組候選基因的表達有明顯的影響(圖6)。然后進行RNA-seq分析,對CTD缺失的后果進行更全面的展示(圖7B-7K)。當比較p53+/-肝臟和相應的p53Δ24/-組織時,274個基因在輻射后3小時有差異表達。GSEA顯示,p53+/-肝臟中表達量增加的基因所富集的GO項與p53Δ24/-肝臟中表達量增加的基因所富集的GO項不同。值得注意的是,p53Δ24/-肝臟中調控增強的基因的GO富集包括 "程序性細胞死亡"(圖7C)。這些差異也可能是由于基礎基因表達的改變。為從進一步分析中排除這些基因,在0 h時對兩種基因型的肝臟進行了比較(圖S7A)。這樣就剩下了241個基因,它們代表了那些基礎表達無顯著統計學差異,但在3 h時調控發生改變的基因(圖7D)。其中,191個基因在p53+/-肝臟中增加,而50個基因在p53Δ24/-組織中增加(圖7E)。因此,有兩類基因:僅在p53+/-肝臟中被激活而在p53Δ/-中未被激活的基因(191個基因)和僅在p53Δ/-肝臟中被激活而在p53+/-中未被激活的基因(50個基因)(圖7E)。對每組基因都進行了檢查,以確認它們確實具有輻射反應性。
第一組中的191個基因在p53+/-肝臟中是靶基因,但在p53Δ/-組織中失去了反應性,其中只有46個基因具有輻射反應性(圖7F)。這些基因似乎是肝臟特異性的,因為大多數基因在胸腺或脾臟中不會隨輻射而改變(圖7G)。在這種情況下,CTD似乎是肝臟特異性基因表達所必需的。與ChIP-seq研究結果(圖1H)結合顯示,這些基因被p53特異性地占據(圖7H)。因此,有一部分靶點是肝臟特異性的(在胸腺或脾臟不受調控),它們的激活依賴于完整的CTD。肝臟特異性與基因在肝臟而非胸腺或脾臟的選擇性占據有關。CTD在這種情況下發揮作用的根本原因可能是通過多種機制,仍有待闡明(圖7L)。
在p53Δ24/-肝臟中顯示調控增強的50個基因中,13個基因在p53Δ/-肝臟中被輻射誘導,而在p53+/-肝臟中沒有被輻射誘導(圖7I)。事實上,其中許多是胸腺和脾臟的靶標(圖7J)。事實上,其中兩個(Rhbdf2、Rxylt1)在胸腺和脾臟中是野生型p53的靶點,但在肝臟中不是(圖7I和7J)。CTD的缺失使這些基因在肝臟中被激活。將這些結果與同基因+/+小鼠的ChIP-seq數據(圖1G)相結合,可以發現所有這些基因都被肝臟p53占據(圖7K)。這表明這些基因與肝臟中的p53相關,但只有在缺失CTD時才能被激活。一種可能的解釋是抑制因子與肝臟中的CTD相關(圖7M)。CTD的缺失將阻止抑制因子的結合,從而導致激活。值得注意的是,有33個基因在基本或輻射后CTD缺失時均表現出差異表達(圖7D)。有趣的是,其中許多基因屬于20個跨組織共享基因(圖2)。其中三個基因(Aen, Eda2r, Zfp365)通過qRT-PCR得到證實。在這些情況下,X-射線照射后出現的激活增強不僅僅是由于基礎表達的增加。
GSEA顯示,這兩組基因(CTD需要表達的基因和CTD抑制表達的基因)具有不同的GO術語(圖7L和7M)。
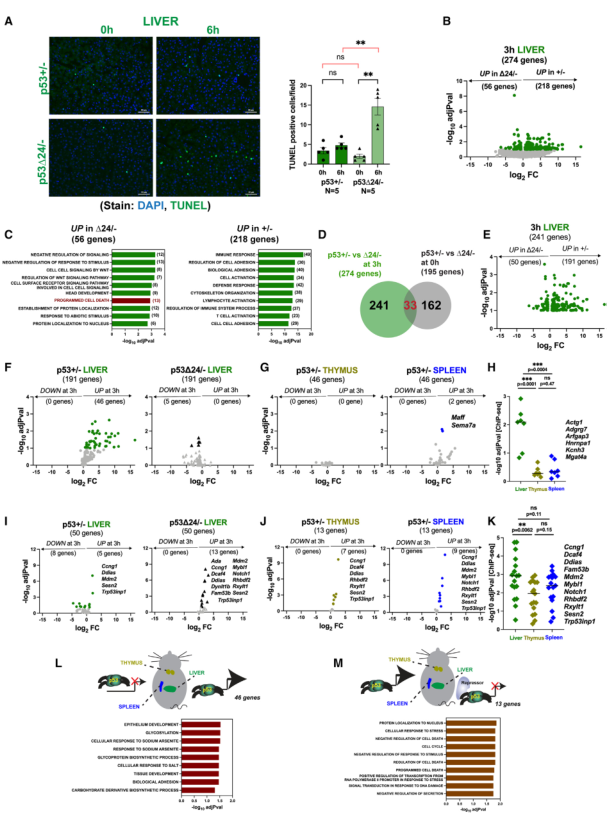
圖7 p53的C端阻止X射線誘導的肝臟凋亡,并決定肝臟特異性基因的表達
總之,如下圖所示,本研究利用RNA-seq和ChIP-seq發現p53輻射反應具有共享性和組織特異性;肝臟p53占據凋亡靶基因但不能調控其表達;p53 C末端抑制肝臟凋亡基因,賦予放射抗性;肝臟特異性基因對輻射反應需要p53 C末端。
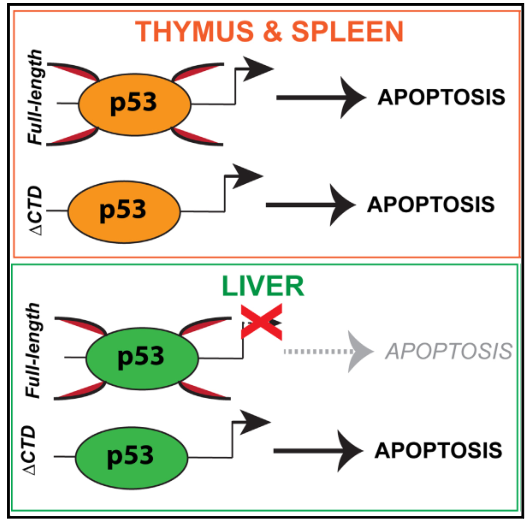
圖8 圖形摘要
實驗方法
小鼠輻射建模和多柔比星處理,RNA提取,qRT-PCR,RNA-seq,ChIP-seq,TUNEL,免疫印跡
參考文獻
Resnick-Silverman L, Zhou R, Campbell MJ, Leibling I, Parsons R, Manfredi JJ. In vivo RNA-seq and ChIP-seq analyses show an obligatory role for the C terminus of p53 in conferring tissue-specific radiation sensitivity. Cell Rep. 2023 Mar 28;42(3):112216. doi: 10.1016/j.celrep.2023.112216. Epub 2023 Mar 15. PMID: 36924496; PMCID: PMC10200257.