






新亞群發高分——CD36+ CAFs的免疫功能
實驗方法:CRISPR-Cas9敲低小鼠和模型構建,單細胞RNA測序和生物信息學分析,慢病毒包裝,ELISA,ChIP,RT-qPCR,多重免疫熒光(mIF)染色,IHC實驗,細胞共培養,脂肪酸或脂蛋白的攝取、脂質過氧化實驗,核質分離實驗,流式細胞術,遷移抑制因子(MIF)染色實驗
肝細胞癌(HCC)是一種具有高度細胞異質性的免疫治療耐藥惡性腫瘤。細胞類型的多樣性以及腫瘤細胞與非腫瘤細胞之間的相互作用仍有待闡明。人類和小鼠HCC腫瘤的單細胞RNA測序顯示癌癥相關成纖維細胞(CAF)的異質性。本研究進一步分析證實CD36+ CAFs通過分泌巨噬細胞遷移抑制因子為HCC提供免疫抑制微環境。
技術路線:

主要實驗結果:
1、單細胞分析揭示人HCC腫瘤的復雜性
對7例原代HCC腫瘤組織和癌旁組織進行單細胞RNA測序(圖1a)。對細胞進行過濾和歸一化后進行主成分分析,使用已知的細胞markers細胞共分為9種類型,包括上皮和腫瘤細胞,B細胞,T細胞,NK細胞,髓源性抑制細胞(MDSCs),單核細胞或巨噬細胞,樹突狀細胞,成纖維細胞,內皮細胞(圖1b和1c)。
2、人HCC中5種CAFs亞型的鑒定和分子特征
α-SMA的IHC染色顯示,80%以上的HCC病例以成纖維細胞激活和積累引起的廣泛肝纖維化為特征(圖1d)。對本研究單細胞RNA測序中的CAFs進行進一步聚類,發現其分為5種亞型(圖1e),它們都高表達經典成纖維細胞markers,然而,不同亞群表型出截然不同的轉錄組特征(圖1f-g)。其中Subcluster 0 CAFs占CAF群體的約40%,并以MYH11、MUSTN1和MCAM等標志性微血管基因為特征(圖1f-g)。因此,將其命名為血管CAFs(vCAFs,vCAFs-c0-MYH11)。GO和KEGG分析顯示vCAFs顯著富集至血管平滑肌收縮和對鈣離子的反應,與它們的微血管特征一致(圖1h-i)。Subcluster 2 CAFs表達低水平α-SMA和高水平ECM特征(圖1f-g)。GO顯示這個亞群與ECM和膠原纖維組織有關(圖1h-i),因此,將其命名為matrix CAFs(mCAFs,mCAFs-c2-LUM)。此外,Subcluster 1 CAFs高表達脂質相關基因(圖1f-g),且富集至ECM,膽固醇代謝,脂肪酸代謝等,提示其可能參與ECM和膽固醇代謝(圖1h-i),因而命名為脂質處理mCAFs(lpmCAFs,lpmCAFs-c1-CD36)。類似的,根據其高表達基因特性和KEGG富集特征,將Subcluster 3 CAFs命名為脂質處理CAFs(lpCAFs,lpCAFs-c1-APOA1);Subcluster 4 CAFs命名為抗原呈遞CAFs(apCAFs,apCAFs-c4-HLA-DRA)。為探索不同病因引起的CAF亞型的異質性,對7個非HBV相關的HCC腫瘤進行了scRNA-seq,發現與HBV-HCC相比,非HBV HCC中CD36+ CAFs和lpCAFs的比例顯著降低(圖1j-l)。最后,使用多重免疫熒光染色證實了人HCC樣本中存在主要CAFs亞群(圖1m)。
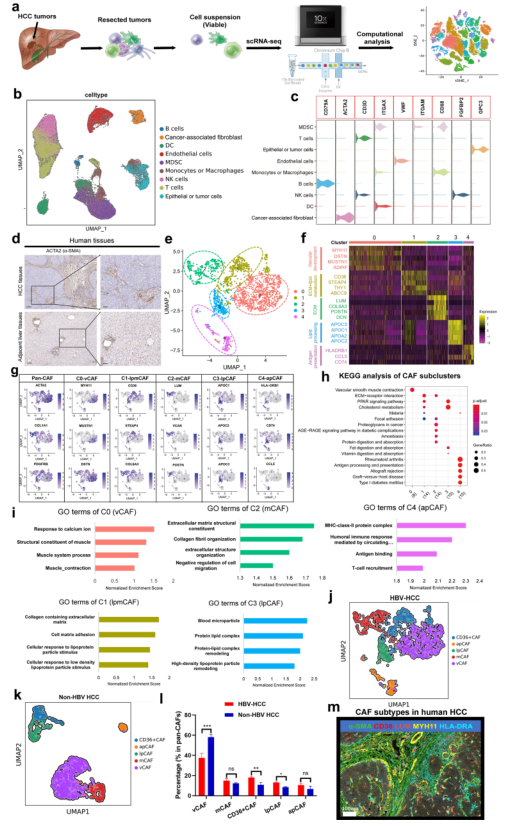
圖1人類HBV相關HCC中不同的成纖維細胞亞群
3、小鼠HCC腫瘤中CAF亞型的單細胞分析概括了人類HCC中發現的亞型
在上述人的HCC樣本證實存在5種CAFs,為進行更深入的CAF表征,將研究擴展到小鼠HCC模型。將px330-sg-p53和CMV-SB13聯合CTNNB1-N90尾靜脈注入6周齡C57BL/6J小鼠,建立CTNNB1N90;Trp53KO肝癌小鼠模型,以模擬肝癌遺傳背景(圖2a)。基于該模型,將7個肝癌小鼠腫瘤分離成單細胞,嚴格過濾后共獲得63977個活細胞,分為10個簇(圖2b)。隨后,其中1746個CAFs聚為7個亞群(圖2c),它們都高表達經典成纖維細胞markers,然而,不同亞群表型出截然不同的轉錄組特征(圖2d-e)。有趣的是,在人樣本鑒定到的5個CAFs亞群也都存在于這小鼠樣本中(圖2c)。CD36+ CAFs和lpCAFs富集了包括ROS和代謝在內的上調通路(圖2f)。最后,使用Myh11、Lum、Acta2、Cd36和H2-Aa等特異性標記物進行mIF染色,以確認主要CAF簇也存在于小鼠HCC組織中(圖2g)。
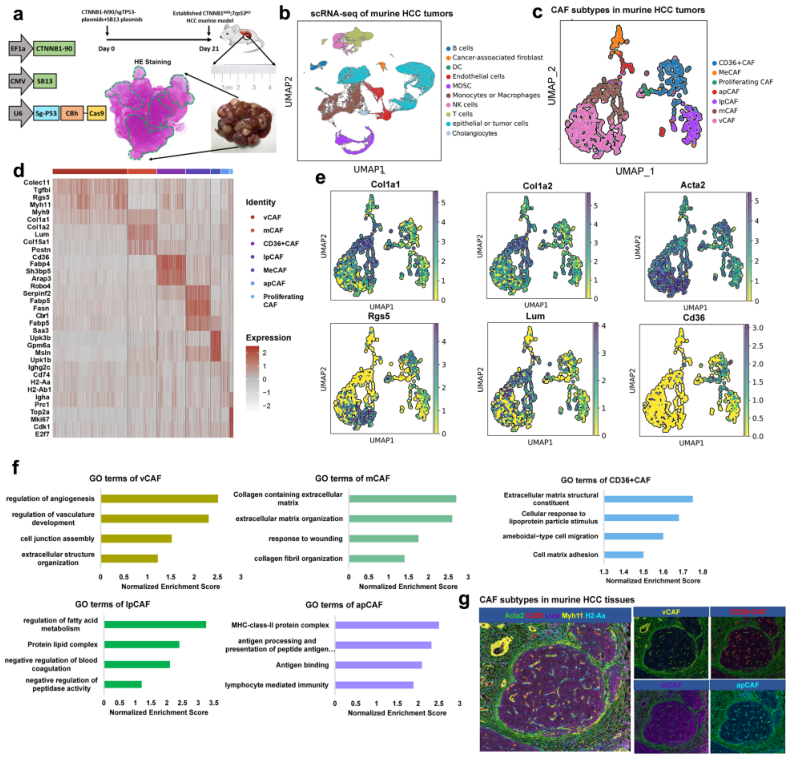
圖2小鼠HCC組織中不同的成纖維細胞亞群
4、CAFs亞型特異性轉錄因子(TFs)和基因調控網絡
接下來,確定TFs及其靶基因調控網絡,以更好地理解CAF亞型如何在遺傳上建立和維持。為此,應用SCENIC21軟件確定了在一種CAF亞型中相對于其他亞型具有高度活性的TFs及其靶點。觀察到vCAFs富集于MEF2C和FOS(圖3a, b),而MEF2C和FOS先前被認為調節新生血管生成。此外,MEF2C的靶基因MYLK、ACTA2和MYH11在vCAFs中表達上調(圖3d)。TWIST1和CREB3L1是mCAFs中最高表達TFs(圖3a, c)。TWIST1是CAF轉分化的關鍵因子。TWIST1靶基因大部分在mCAFs中上調(圖3d)。lpmCAFs (CD36+ CAFs)顯示CEBPD的高表達(圖3a, c)。脂蛋白脂肪酶LPL靶基因在lpmCAFs中上調表達(圖3d)。除了CEBPD,lpCAFs還高表達CEBPA和MAFB(圖3a, c),它們是已知TF參與促進LDL代謝轉錄程序。apCAFs高表達IKZF1和RUNX3(圖3a, c),它們與巨噬細胞極化和T細胞招募有關。為檢測在這些小鼠CAF亞群中活躍的主調控因子,進行SCENIC分析,結果顯示Mef2c和Foxf1是小鼠vCAFs中活躍的TF基因,與人類vCAFs相似(圖3e,f)。
5、CD36+ CAFs與MDSCs呈正相關,在HCC腫瘤中與MDSCs接近
接下來探索異質性CAF與TME中其他細胞之間的特異性串擾機制。在所有已知的配體-受體對中,MIF信號通路以MIF配體及其CD74/CXCR4受體為主。有趣的是,lpmCAFs和lpCAFs高表達CD36(圖1g),CD36是作用于MDSCs的MIF配體的最主要來源。因此,作者使用特定的表面標記在lpmCAFs (c1-mCAF-CD36)和lpCAFs中鑒定CD36,并發現它們都在脂肪生成通路中富集(圖3g)。mIF實驗表明,與癌旁組織相比,CD36+ CAFs在小鼠和人的腫瘤組織中特異性浸潤(圖3h, i)。隨后研究了CD36+ CAFs與TME內效應T細胞MDSCs之間是否存在相關性。scRNA-seq數據顯示CD36+ CAFs與MDSCs呈正相關,但和效應T細胞呈負相關(圖3j),該結果被臨床切片組織染色證實(圖3k-l)。此外,對12例HCC腫瘤進行單細胞核測序,發現CD36+ CAFs的數量與MDSCs的數量呈正相關(圖3m)。mIF染色顯示CD36+ CAFs比CD36- CAFs更接近MDSCs(圖3n)。總之,這些結果表明CD36+ CAFs可能賦予HCC有效的免疫抑制作用。
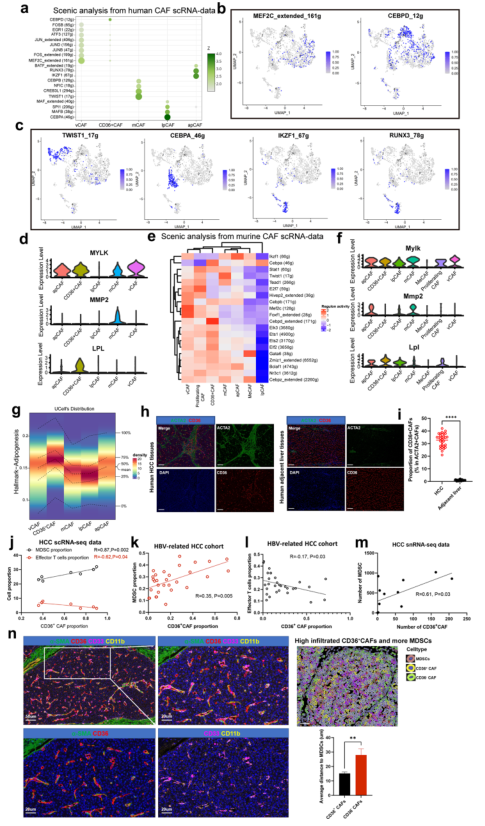
圖3 CAFs的特異性TF以及CAFs和MDSCs之間的相互作用
6、CD36在CD36+ CAFs中通過脂質過氧化/p38/CEBPs軸介導OxLDL攝取促進MIF表達
利用圖4a中的流式細胞術策略分離CD36+ CAFs,此外,其分離的CAFs細胞類型比例和小鼠即人樣本中的分布類似(圖4b-c)。隨后測試不同類型CAFs產生MIF的水平。結果顯示CD36+ CAFs產生的MIF水平顯著高于其它類型(圖4d,e)。基于這些結果,作者將重點放在CD36+ CAFs的這個亞簇上進行進一步研究。WB和ELISA證實當在CD36+ CAFs細胞中沉默CD36(CD36kd CAFs)后MIF的表達和分泌顯著減少(圖4f)。此外,與前文單細胞測序結果的信號通路分析類似,作者發現CD36+ CAFs中脂質過氧化相關基因的表達顯著高于CD36kd CAFs(圖4h)。使用熒光偶聯OxLDL和流式細胞術,發現CD36+ CAFs比CD36kd CAFs有更高的OxLDL攝取率(圖4i, j)。CD36+ CAFs的脂質過氧化也顯著高于CD36kd CAFs(圖4k)。接下來確定OxLDL是否會增加CD36+ CAFs中的脂質過氧化,結果顯示OxLDL以劑量依賴的方式增強CD36+ CAFs的脂質過氧化作用,而LDL則沒有(圖4l)。此外,通過檢測CD36+ CAFs中p38的磷酸化來檢測OxLDL是否可以激活p38,結果發現OxLDL誘導CD36+ CAFs的p38磷酸化,但在CD36kd CAFs中程度較低,并且添加Toco或SSO降低了OxLDL誘導的p38磷酸化(圖4m),并且p38磷酸化在CD36+ CAFs中的富集顯著高于CD36敲除或陰性CAFs細胞(圖4n)。這些表明OxLDL通過CD36和脂質過氧化促進p38激活。在OxLDL存在時,p38抑制部分挽救了MIF的分泌(圖4o),表明OxLDL部分通過p38激活促進MIF的分泌。
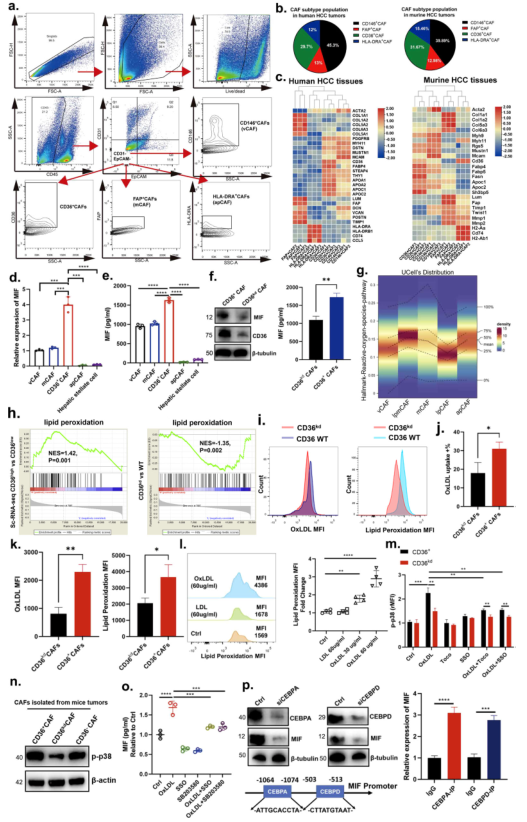
圖4CD36在CD36+ CAF中通過脂質過氧化/p38/CEBPs軸介導OxLDL攝取促進MIF表達
7、CD36+ CAF來源的MIF通過激活IL-6/ STAT3增強MDSCs促進免疫抑制性TME和腫瘤干性的能力
為確定來自腫瘤細胞或CD36+ CAFs的MIF是否介導CD33+ MDSC擴增的調節,從人HBV相關或小鼠HCC腫瘤中分離出人或鼠CD36+ CAFs。采用CD36+ CAFs條件培養基(CD36+ CAFs-CM)、CD36 - CAFs、CD36kd CAFs-CM、CD36+ CAFs +MIF抑制(ISO-1)或CD74阻斷處理MDSC前體細胞,檢測CD33+ MDSC比例(圖5a, b)。CD36+ CAFs-CM,MIF可增加CD33+ MDSCs的數量;與CD36kd CAFs-CM類似,CD36+ CAFs +ISO-1和CD36+ CAFs +CD74阻斷表現出良好的抑制效應在CD33+ MDSC擴增時(圖5c)。表明MIF分泌是CD36+ CAFs調節MDSC擴張的關鍵中介。
在免疫正常小鼠的原位肝癌模型中,發現注射從小鼠肝癌腫瘤中分離的CD36+ CAFs顯著促進肝癌腫瘤細胞在小鼠肝臟中的生長或轉移,而這種促進作用被特異性敲低CD36,抑制MIF或去除Gr-1+ MDSCs所抑制(圖5d, e)。此外,注射CD36+ CAFs顯著增加Treg的浸潤,但減少腫瘤中效應CD8+ T細胞,這種作用同樣被特異性敲低CD36,抑制MIF或去除Gr-1+ MDSCs所抑制(圖5f, g)。
為研究CD36+CAFs是否能通過iNOS信號調節MDSCs以增強其免疫抑制能力,從原代CD36+CAFs、CD33+MDSCs中收集CM,或分別從CD36+CAFs、CD36kd CAFs、CD36+ CAFs +CD74的CM預處理的CD33+MDSCs的培養物中收集CM,評估iNOS的水平。結果顯示CD36+CAFs-MDSC-CM,而不是CD36kd CAF-MDSC-CM或來自CD74或MDSC阻斷的細胞的CM,極大地促進了MDSCs的iNOS的分泌(圖5h)。此外,共培養實驗顯示,與MDSC-WT組相比,CD36+ CAFs+MDSC組的CD69+、IFNG+CD8+T細胞的比例明顯下調,但Tregs和PD1+CD8+T細胞的比例上調,而CD36+ CAFs+MDSC+ISO-1或CD74阻斷引起的影響輕微(圖5i, j)。表明CD36+ CAFs發揮免疫抑制作用是以CD36依賴的方式通過MIF/CD74/iNOS軸。
最后,為探索CD36+CAF衍生的MIF影響MDSCs的分子機制,對MDSC-MIF組和MDSC組進行轉錄組測序。結果顯示,NF-κB信號通路被排在首位,其被報道可調節MDSCs的細胞因子分泌,在用CD36+CAF衍生的MIF預處理的MDSCs中被激活(圖5k, l)。此外,CD36+CAF處理的MDSCs中p65磷酸化顯著增加(圖5m),iNOS和IL-6的分泌也顯著增加(圖5n)。這些表明CD36+CAF促進MDSCs是通過IL-6介導的。
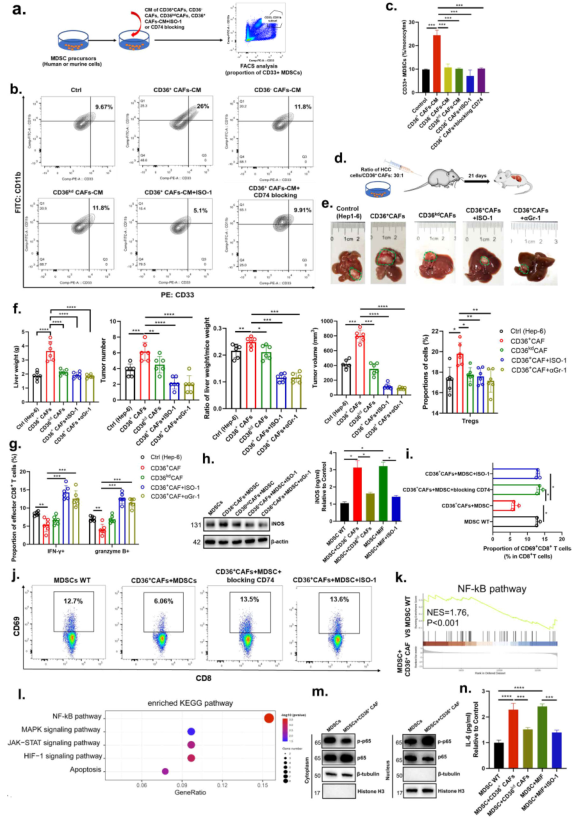
圖5 CD36+CAF來源的MIF通過激活IL-6/ STAT3增強MDSCs促進免疫抑制性TME和腫瘤干性的能力
8、在小鼠模型中靶向CD36協同免疫治療
為進一步研究CD36+和MIF+CAFs是否在HCC啟動中發揮作用,建立Cd36(Acta2Cre;Cd36fl/fl)和MIF(Acta2Cre;Miffl/fl)條件性敲除的6周齡小鼠,確定Cd36或MIF在體內被敲除時,HCC腫瘤負擔和MDSCs的比例明顯減少(圖6a-d),這表明CD36+CAFs參與HCC的啟動。所以作者猜想CD36+CAFs可能與HCC患者的免疫治療有關。因此,首先調查了離體的HCC組織中的CD36+CAFs,這些患者接受了新輔助抗PD-1治療。結果顯示,抗PD-1反應組的CD36和α-SMA共表達低于無反應組,表明低數量的CD36+CAFs可以預測更好的HCC免疫治療反應(圖6e,f)。然后,我們在C57/BJ6 CTNNB1N90;Trp53KO HCC模型和作者建立的抗PD1耐藥HCC中探索了單藥(CD36抑制劑或抗PD-1療法)和聯合治療策略(CD36抑制和抗PD-1療法)的療效。發現,聯合治療在這些HCC小鼠模型中表現出明顯的抗腫瘤療效(圖6g-k),并改變了抗腫瘤免疫的免疫格局,Tregs和MDSCs的比例下降,IFN-γ+和顆粒酶B+CD8+T細胞的比例增加(圖6l-m)。因此,腫瘤中低數量的CD36+CAFs可能預示著HCC有更好的免疫治療反應,用SSO靶向CD36可以用來協同提高免疫治療的效果。
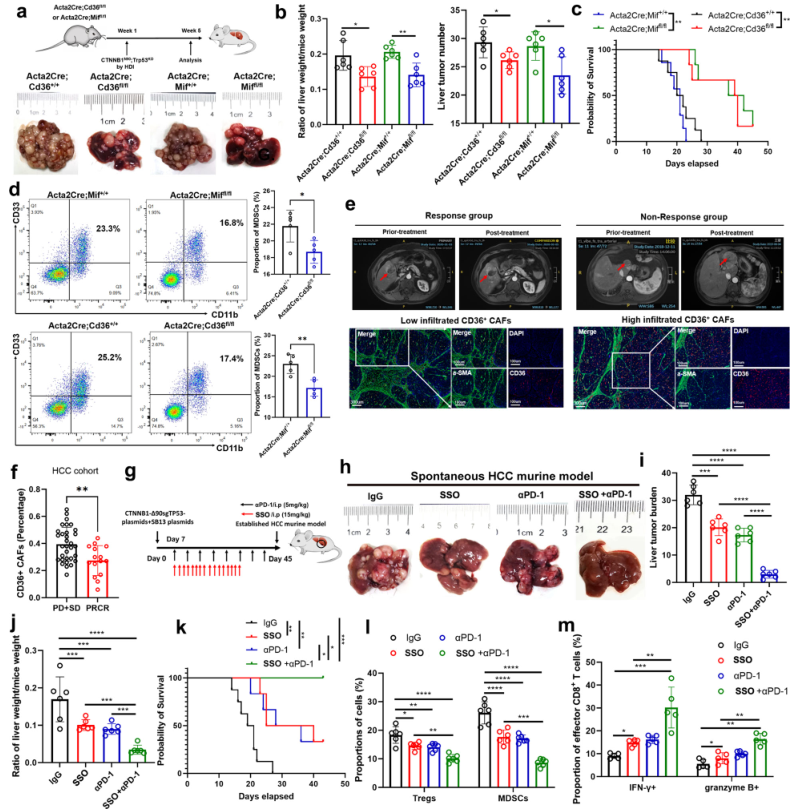
圖6CD36+ CAFs預測肝癌免疫治療的療效和靶向MIF與肝癌小鼠免疫治療的協同作用
參考文獻:
Zhu, GQ., Tang, Z., Huang, R. et al. CD36+ cancer-associated fibroblasts provide immunosuppressive microenvironment for hepatocellular carcinoma via secretion of macrophage migration inhibitory factor. Cell Discov 9, 25 (2023). https://doi.org/10.1038/s41421-023-00529-z