






ALKBH3依賴的Aurora A mRNA的m1A去甲基化抑制纖毛生成
初級纖毛是觸角狀的亞細胞結構,作為信號平臺,調節許多細胞過程和胚胎發育。m1A RNA修飾在RNA代謝和基因表達中起關鍵作用;然而,m1A修飾的生理功能在很大程度上仍不清楚。我們發現m1A去甲基化酶ALKBH3通過其去甲基化活性顯著抑制哺乳動物細胞纖毛發生。機制上,ALKBH3去除纖毛發生的主抑制因子Aurora A mRNA上的m1A位點。ALKBH3的缺失增強了Aurora A mRNA的衰減并抑制其翻譯。此外,在斑馬魚胚胎中,ALKBH3突變體表現出纖毛缺陷,包括彎曲體、心包水腫、異常耳石和前腎管擴張,野生型ALKBH3顯著拯救了這些缺陷,但其催化失活突變體卻不能。在脊椎動物細胞和胚胎中,由ALKBH3耗盡引起的纖毛缺陷也被Aurora A mRNA的異位表達顯著逆轉。總之,我們的數據表明,依賴于ALKBH3的m1A去甲基化在Aurora A mRNA的調控中起著至關重要的作用,這對脊椎動物纖毛發生和纖毛相關的發育事件至關重要。本文于2022年3月發表于Cell Discovery (IF=38.079)上。
技術路線:

主要研究結果:
(1) ALKBH3通過依賴m1A的方式抑制纖毛發生
為了研究m1A RNA修飾在纖毛發生中的作用,我們首先在正常條件下使用小干擾RNA (siRNA)對人類RPE-1細胞中編碼m1A調節蛋白的靶向基因進行了功能篩選。TRMT6、TRMT61A或TRMT61B的缺失對RPE-1細胞纖毛發生沒有顯著影響(補充圖未展示)。用兩種siRNA中的一種沉默ALKBH1或TRMT10C對纖毛比例有影響。重要的是,用兩種不同的siRNA敲除ALKBH3顯著增加了纖毛細胞的百分比,但不影響纖毛的長度(圖1a-d)。此外,ALKBH3-FLAG蛋白的異位表達顯著逆轉了ALKBH3敲除細胞中增強纖毛的作用(圖1e-g)。為了確認ALKBH3在纖毛中的功能,我們外源表達了ALKBH3-FLAG,發現強制表達ALKBH3- FLAG顯著減少了血清饑餓處理的RPE-1細胞的纖毛發生(圖1h-j)。
由于ALKBH3在mRNA上的去甲基化活性被D193A的突變所抑制,我們制備了一個催化不活躍的ALKBH3突變體(ALKBH3D193A),以研究ALKBH3在纖毛發生中的調控是否依賴于它的去甲基化活性。通過LC-MS/MS對RPE-1細胞中m1A mRNA修飾的定量分析,證實我們構建的ALKBH3-D193A結構確實是催化活性不高的(補充圖未展示)。野生型ALKBH3-FLAG的異位表達顯著恢復了ALKBH3耗盡細胞的纖毛表型,但在RPE-1細胞中,突變型ALKBH3-D193A-FLAG的異位表達無效(圖1k-m)。此外,過表達的ALKBH3-D193A突變體對血清缺失的RPE-1細胞的纖毛沒有顯著影響(補充圖未展示)。總之,這些數據表明m1A RNA去甲基化酶ALKBH3在哺乳動物細胞中起著纖毛發生的負調控作用。
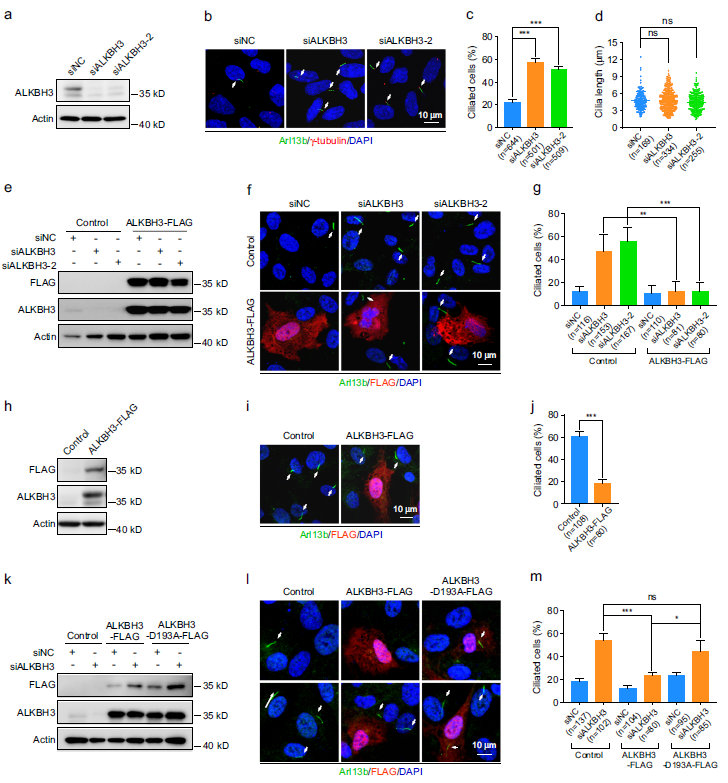
圖1:ALKBH3通過其m1A去甲基化活性抑制纖毛發生。
(2) 敲低ALKBH3導致Aurora A mRNA的m1A水平升高
為了探索ALKBH3在纖毛發生中的分子機制,我們試圖確定被ALKBH3靶向的可能的mRNAs。首先,我們系統地分析了已發表的HEK-293T細胞中m1A-ID-seq和m1A-quant-seq的數據集,獲得了38個潛在的m1A修飾的轉錄本。這些潛在的m1A修飾的轉錄本進一步與來自CiliaCarta的纖毛蟲相關基因和在缺乏ALKBH3的RPE-1細胞中差異表達的RNA-seq基因重疊。結果,在這四個數據集中,只有Aurora A,一個主要的負調節因子,是重疊的(圖2a)。為了研究Aurora A mRNA上是否存在m1A位點,我們進行了基因特異性甲基化RNA免疫沉淀qPCR試驗。數據顯示,與對照細胞相比,在缺乏ALKBH3的細胞中,Aurora A mRNA的m1A豐度顯著增加(圖2b),表明Aurora A mRNA可能包含m1A修飾。
為了確定Aurora A mRNA上的精確m1A位點,我們仔細分析了來自野生型和ALKBH3敲除HEK-293T細胞中的m1A-ID-seq數據集,發現在Aurora A mRNA的翻譯起始區附近有潛在的m1A峰(圖2c)。考慮到RT-1306是一種進化的HIV逆轉錄酶,能夠誘導RNAs上m1A位點的突變,我們使用RT-1306來識別Aurora A mRNA上的m1A位點。我們從對照細胞和ALKBH3缺失細胞的cDNAs中獲得的測序數據顯示,在Aurora A mRNA翻譯起始位點附近的編碼序列區存在3個m1A位點(圖2d)。此外,我們還將RT-1306擴增的上述cDNAs進行TA克隆,并選取單個菌落進行測序分析。結果顯示,在缺乏ALKBH3的細胞中,這些m1A位點的突變率顯著增加(圖2e)。綜上所述,這些觀察結果表明,ALKBH3催化Aurora A mRNA上m1A位點的去甲基化。
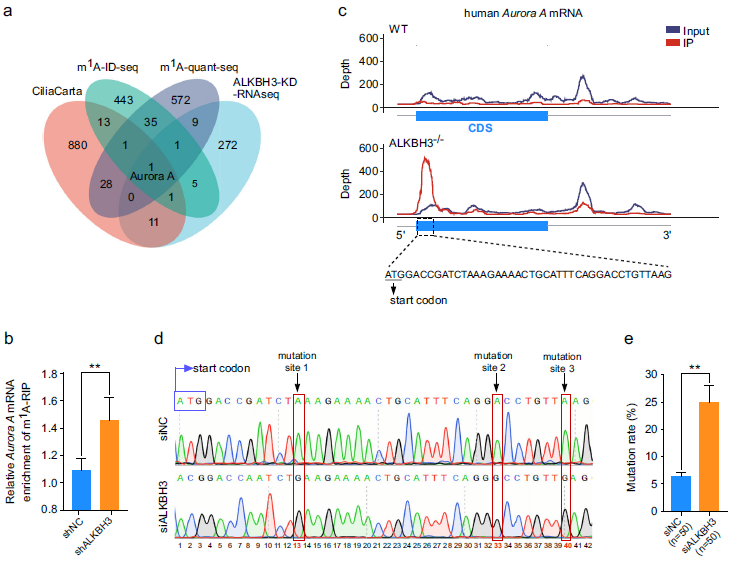
圖2:ALKBH3去除Aurora A mRNA上的m1A位點。
(3) ALKBH3促進Aurora A mRNA的穩定和翻譯
由于mRNA的m1A修飾被報道影響mRNA的代謝過程,包括mRNA的穩定性和翻譯,我們首先研究了ALKBH3是否調節Aurora A的mRNA水平。結果顯示,敲低ALKBH3顯著降低了RPE-1細胞中Aurora A mRNA的水平(圖3a,b)。進一步的實驗表明,當細胞中使用放線菌素D暫停轉錄時,在缺乏ALKBH3的細胞中,Aurora A mRNA的衰減率也顯著增強(圖3c)。接下來,我們確定了Aurora A mRNA的RNA翻譯效率是否受ALKBH3的影響。Western分析表明,ALKBH3的缺失顯著降低了Aurora A蛋白水平,而異位表達的ALKBH3有效逆轉了這一變化,但突變體ALKBH3-D193A卻沒有改變(圖3d-g)。此外,RPE-1細胞中的多聚體分析顯示,穩定敲除ALKBH3會導致高分子量(HMW)多聚體部分的Aurora A mRNA豐度降低,而這些多聚體通常具有較高的翻譯效率(圖3h,i),這意味著ALKBH3的缺失可能會抑制Aurora A mRNA的翻譯。總的來說,這些數據表明,ALKBH3對Aurora A mRNA的穩定性和翻譯的調節至關重要。
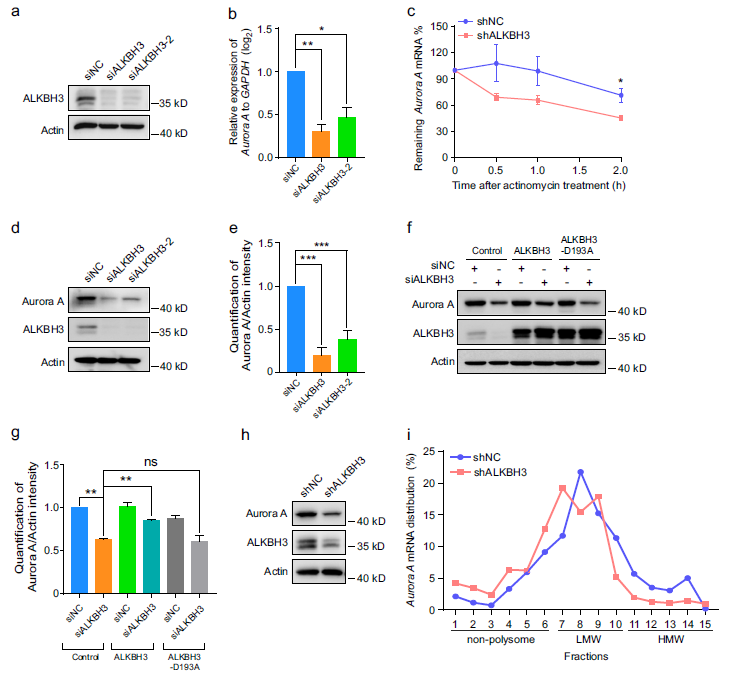
圖3:敲低ALKBH3會降低Aurora A mRNA的穩定性和翻譯能力。
(4) ALKBH3通過Aurora A抑制纖毛發生
考慮到Aurora A是促進纖毛分解的關鍵調控因子,我們推斷ALKBH3可能影響纖毛分解。為了驗證這一點,我們用血清饑餓處理RPE-1細胞24小時以誘導纖毛,然后用血清再刺激24小時以觸發纖毛拆卸(圖4a)。在此條件下,我們觀察到對照組RPE-1細胞在血清刺激24小時后纖毛顯著減少(圖4b-d)。然而,缺乏ALKBH3的細胞明顯抵抗血清誘導的纖毛拆卸。因此,這些數據表明,ALKBH3具有調節纖毛拆卸的功能。我們的數據顯示,Aurora A的總水平在血清刺激后2小時略有上升,在18小時和24小時達到峰值(圖4e)。我們還發現在血清饑餓/再刺激期間,ALKBH3與Aurora A的表達模式相似(圖4e)。值得注意的是,在整個血清刺激過程中,缺乏ALKBH3的RPE-1細胞中的Aurora A蛋白水平明顯降低(圖4f),這意味著ALKBH3對Aurora A表達的調控可能發生在纖毛周期中。
免疫熒光實驗數據顯示,在缺乏ALKBH3的細胞中,Aurora A和磷酸化的Aurora A水平都明顯降低(補充圖未展示)。進一步的結果表明,在缺乏ALKBH3的細胞中,中心體上Aurora A和磷酸化的Aurora A的熒光強度顯著降低(補充圖未展示)。中心體上Aurora A和磷酸化的Aurora A的細胞比例也顯著降低(補充圖未展示)。這些觀察結果表明,ALKBH3的消耗降低了中心體Aurora A和磷酸化Aurora A的水平。此外,Aurora A的沉默顯著促進了RPE-1細胞的纖毛發生(補充圖未展示)。拯救實驗表明,外源表達GFP-Aurora A蛋白能夠顯著逆轉由ALKBH3敲低引起的增強的纖毛(圖4g-i)。這些數據共同表明,ALKBH3通過促進Aurora A的表達來抑制纖毛。
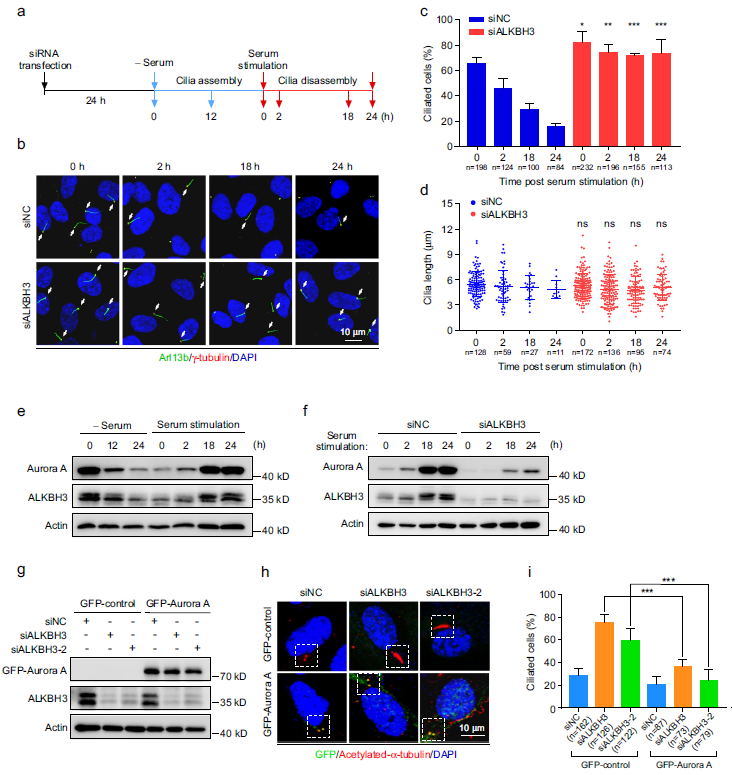
圖4:Aurora A的異位表達逆轉了ALKBH3缺失細胞的纖毛缺陷。
(5) ALKBH3缺乏促進纖毛誘導細胞周期阻滯
為了探索缺乏ALKBH3的細胞中增強的纖毛是否與細胞周期阻滯相關,我們首先檢測了無血清饑餓的RPE-1細胞中缺乏ALKBH3對細胞周期進展的影響。FACS分析顯示,ALKBH3缺失增加了G0/G1期細胞的百分比(補充圖未展示)。免疫印跡分析顯示,在ALKBH3缺失的細胞中,細胞周期標志物cyclin A減少,表明在ALKBH3敲低的細胞中,S期或G2/M期細胞減少(補充圖未展示)。進一步的EdU-標記實驗顯示,當ALKBH3耗盡時,EdU-結合細胞的百分比顯著下降(補充圖未展示)。這些數據表明,ALKBH3的耗盡導致細胞周期在G0/G1期停止。然后,我們研究了由ALKBH3耗竭引起的G0/G1期細胞周期阻滯是否由其增強的纖毛介導。來自FACS譜、細胞周期標記和EdU合并的數據顯示,IFT20耗盡誘導的纖毛脫落顯著逆轉了ALKBH3耗盡細胞中G0/G1阻滯(補充圖未展示),表明ALKBH3耗盡以纖毛依賴的方式誘導G0/G1阻滯。
接下來,我們研究了Aurora A敲除是否也會導致RPE-1細胞中纖毛依賴的細胞周期阻滯。Aurora A的沉默會導致細胞周期在G0/G1期停止,同時IFT20的耗盡也會顯著逆轉(補充圖未展示)。總的來說,這些結果表明,無論是ALKBH3還是Aurora A的缺失都會導致RPE-1的纖毛增強,這可能隨后導致細胞周期在G0/G1期停止。
(6) 斑馬魚ALKBH3的分子特征
為了探索ALKBH3在脊椎動物發育過程中纖毛發生中的作用,我們首先克隆了斑馬魚ALKBH3(圖5a)。生物信息學分析顯示,ALKBH3編碼一個推導出的282 aa蛋白,具有保守的2OG-Fe(II)加氧酶超家族域,這意味著斑馬魚的ALKBH3也可能具有去甲基化活性(圖5b)。比對分析表明斑馬魚ALKBH3與人類ALKBH3具有高度同源性(圖5c)。然后,我們檢測了斑馬魚ALKBH3在胚胎發育過程中的時間表達,發現ALKBH3 mRNA由母體沉積,并在整個早期斑馬魚胚胎階段普遍表達(圖5d)。全安裝原位雜交顯示,ALKBH3 mRNA由母體提供,在48 hpf(受精后小時)到72 hpf時在大腦中富集,在96 hpf時下降到低水平(圖5e)。
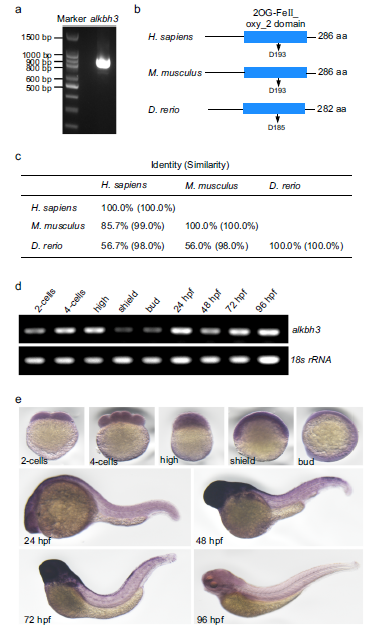
圖5:斑馬魚ALKBH3基因的分子特征。
(7) ALKBH3突變體在斑馬魚胚胎中顯示纖毛表型
為了研究ALKBH3是否影響斑馬魚的絨毛發生和胚胎發育,我們設計了反義嗎啉代寡核苷酸(ALKBH3 MO)來阻斷ALKBH3 mRNA的翻譯。ALKBH3突變體在斑馬魚胚胎中顯示出纖毛表型,包括身體彎曲、心包水腫、耳囊泡內異常耳石和前腎管擴張(圖6a-d)。重要的是,通過與ALKBH3 mRNA共注射,ALKBH3突變體中的這些缺陷被顯著逆轉,但根據氨基酸序列的進化保守分析,預測到催化活性不強的ALKBH3突變體的ALKBH3-D185A mRNA沒有被逆轉(圖5b)。進一步的觀察表明,在28 hpf時,與對照變體相比,ALKBH3突變體的前腎管纖毛明顯有缺陷,異位表達的ALKBH3 mRNA而不是異位表達的ALKBH3-D185A mRNA也顯著挽救了纖毛(圖6e,f)。總之,這些數據表明,ALKBH3可能通過其在斑馬魚體內的去甲基化活性在纖毛相關的發育過程中發揮關鍵作用。
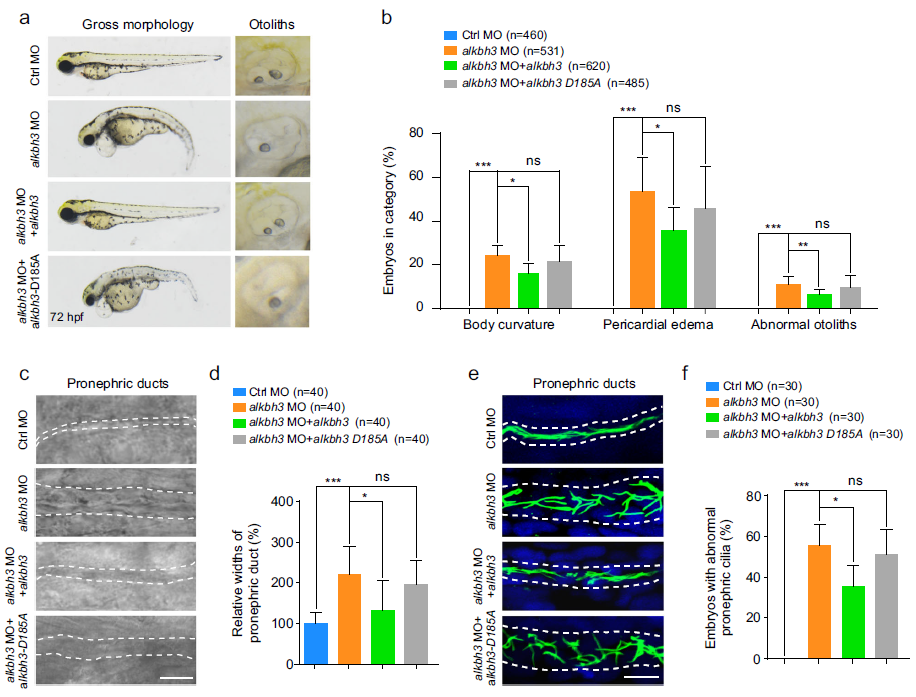
圖6:斑馬魚ALKBH3突變體表現出纖毛表型。
(8) Aurora A的異位表達逆轉了ALKBH3突變體的纖毛缺陷
為了確定Aurora A是否參與斑馬魚胚胎發育過程中ALKBH3對纖毛相關事件的調控,我們檢測了ALKBH3突變體中Aurora A mRNA的表達水平,發現Aurora A mRNA在缺乏ALKBH3的胚胎中顯著降低(圖7a),這與我們從哺乳動物細胞中獲得的數據一致(圖3b)。此外,共注射Aurora A mRNA能夠顯著逆轉ALKBH3突變體的纖毛表型(圖7b-e)。其他數據顯示,異位表達的Aurora A mRNA顯著挽救了ALKBH3敲除胚胎前腎導管的異常纖毛(圖7f,g)。總之,這些結果表明,ALKBH3可能通過調節脊椎動物胚胎發生中的Aurora A mRNA來影響纖毛相關的發育事件。
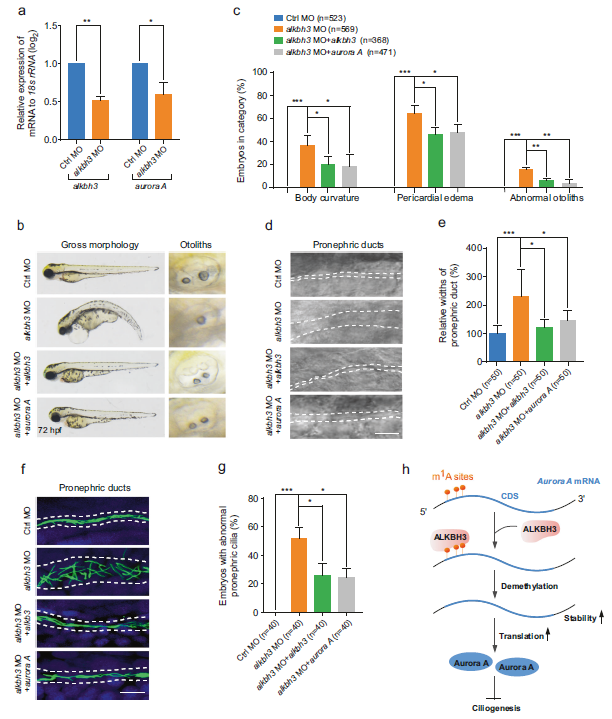
圖7:Aurora A的異位表達挽救了ALKBH3形態的纖毛缺陷。
結論:
我們的研究揭示了m1A去甲基化酶ALKBH3在脊椎動物纖毛發生和胚胎發育中先前未被描述的作用。ALKBH3在翻譯起始位點附近的編碼序列區去除Aurora A mRNA的m1A修飾,抑制Aurora A mRNA的衰變并促進其mRNA的翻譯,從而維持Aurora A蛋白的豐度以抑制纖毛發生(圖7h)。
參考文獻:
Kuang, W., Jin, H., Yang, F., Chen, X., Liu, J., Li, T., Chang, Y., Liu, M., Xu, Z., Huo, C., Yan, X., Yang, Y., Liu, W., Shu, Q., Xie, S., & Zhou, T. (2022). ALKBH3-dependent m1A demethylation of Aurora A mRNA inhibits ciliogenesis. Cell discovery, 8(1), 25. https://doi.org/10.1038/s41421-022-00385-3.